Organelle Adaptations in Plasmodium: The Targets for Malaria Treatments
Some organelle adaptations that support survival in the malaria parasite, Plasmodium, inside the life cycle in vertebrates and mosquitoes include the endoplasmic reticulum, mitochondrion, and apicoplast. This highly unfolded endoplasmic reticulum supports high protein synthesis for rapid parasite growth and replication. The mitochondrion plays a crucial role in this parasite, driving energy production and regulating metabolism. The apicoplast is a relict plastid from secondary symbiosis from a red algalderived source, essential for lipid synthesis, isoprenoid production, and fatty acid elongation. Providing metabolites that are essential and not available from the host. Studies on these organelles may lead to new therapies against diseases like malaria and help resolve global health problems
Abbreviations
FAS: Fatty Acid Synthase; GPI: Glycophosphatidylinositol.
Editorial
Some organelle adaptations that support survival in the malaria parasite, Plasmodium, inside the life cycle in vertebrates and mosquitoes include the endoplasmic reticulum, mitochondrion, and apicoplast. This highly unfolded endoplasmic reticulum supports high protein synthesis for rapid parasite growth and replication. The mitochondrion plays a crucial role in this parasite, driving energy production and regulating metabolism. The apicoplast is a relict plastid from secondary symbiosis from a red algal- derived source, essential for lipid synthesis, isoprenoid production, and fatty acid elongation. Providing metabolites that are essential and not available from the host. Studies on these organelles may lead to new therapies against diseases like malaria and help resolve global health problems.
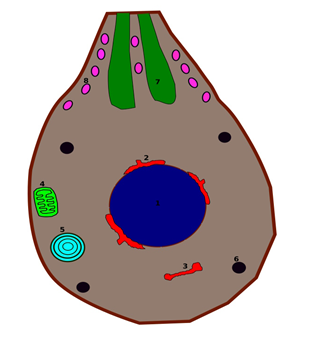
These apicomplexan parasites, especially Plasmodium, undergo dynamic transformations of organelles such as the apicoplast and mitochondria at their critical cycle stage. These organelles have developed some unique adaptations necessary for their parasitic way of life. The apicoplast, secondarily acquired from an ancestral chloroplast, is associated with various metabolic processes such as fatty acid synthesis, production of isoprenoid precursors, and biosynthesis of heme. It is essential for protein targeting and trafficking, and disruption of the pathways proves fatal for the parasite. The apicoplast function makes it a premier drug target, and the metabolic pathway inhibitors show the prospect of disrupting the parasite growth (Figure 1) [1, 2].
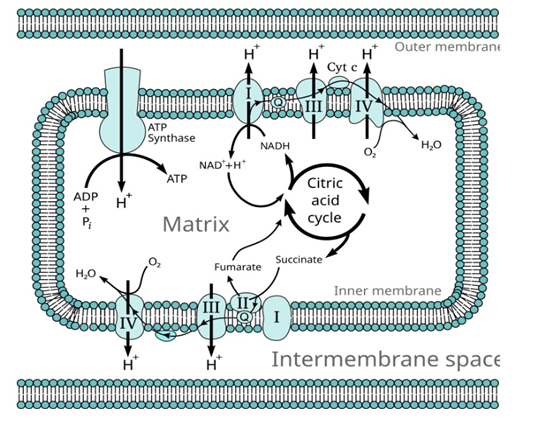
The mitochondrion of apicomplexan parasites has developed some drastic modifications compared to other eukaryotes to accommodate their parasitic style of life. Its respiratory chain is essential for energy production, metabolite synthesis, and redox balance. Unlike typical mitochondria, apicomplexan mitochondria have expanded respiratory complexes and unique electron transport components that have evolved to meet the parasite’s metabolic needs. The mitochondrial respiratory chain is a drug target. Antimalarial drugs such as atovaquone disrupt ATP synthesis by inhibiting the cytochrome bc1 complex, an action that finally results in parasite death. These adaptations have helped develop selective therapies that minimize harm to the host (Figure 2) [3].
The schizont stage underlines the evolutionary adaptability of apicoplast and mitochondrion organelles in an apicomplexan parasite. Such adaptations enable insight into how ancient organelles can be adapted for new functions, affording a better understanding of organelle biogenesis, protein trafficking, and the regulation of metabolism in eukaryotes. The research in this phase will further our knowledge of the interplay between organelle function and parasite biology on apicoplast protein import mechanisms, discovery of new metabolic pathways, and mitochondrial biogenesis. Such knowledge is bound to assist in developing therapies targeted at parasitic diseases [4].
In Plasmodium, the apicoplast is a non-photosynthetic organelle enveloped by four membranes. It is essential for metabolic processes such as lipid biosynthesis and the Type II Fatty Acid Synthase (FAS) pathway. These functions are features of parasite growth and invasion into the host. Because the apicoplast is a prokaryotic-like organelle containing unique metabolic pathways not present in humans, it is an attractive target for anti-malarial drugs. Yet, the parasite survives an initial drug exposure since only completely disabled apicoplast functions result in a parasite-affecting consequence in later replication cycles. Recent studies have investigated the apicoplast synthesis of isoprenoid precursors, essential to major cellular processes such as glycophosphatidylinositol (GPI) anchor biosynthesis. Apicoplast disruption blocks merozoite maturation and its release from erythrocytes. Since then, the interruption has made the asexual phase of the parasite come to an end. These
analyses brought forward the importance of an apicoplast as part of the therapeutic relevance given. Much work is underway to identify quicker-acting drugs; the apicoplast inhibitors hold great promise for new advances in malaria control (Figure 3) [5, 6].
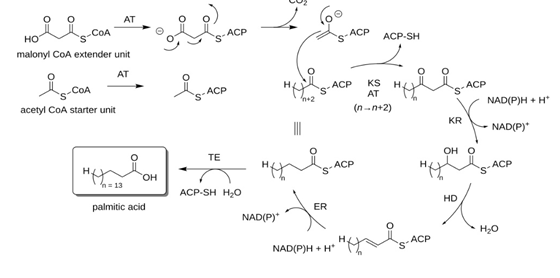
The apicoplast is a unique plastid found in apicomplexan parasites like Plasmodium species. It is essential for parasite survival because it hosts metabolic pathways. This secondarily endosymbiotic organelle has a complex, multilayered membrane system that requires transit peptides to import proteins critical to its function. Because it affects protein transport into apicoplast function, disruption of the former type abrogates biosynthetic function, making apicoplast a prime target of anti-malarial therapies. Indeed, drugs that implicate inhibition of isoprenoid precursor biosynthesis, such as fosmidomycin, show great promise. The delayed- death phenotype that some parasites die in subsequent generations following apicoplast disruption underscores the organelle’s critical role and vulnerability. Continued research into apicoplast biology provides a basis for the innovative pathways for effective and selective treatments against malaria and other diseases caused by apicomplexans [7, 8].
Organellar adaptations of the apicoplast and mitochondrion in Plasmodium underpin the parasite’s survival and related pathogenicity. These features add immense knowledge of the biology behind the parasite, with privileged opportunities for drug development targeted at them. Additional complexity faces the field of drug design, such as the phenotypic effect of delay of death on drugs interfering with the apicoplast and optimization of mitochondrial inhibitors with higher potency that may develop into more efficacious modalities of treatments against malaria infection [9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39].
Acknowledgments
The author thinks Prof. Marc Gentilini and Prof. Martin Danis (Sorbonne University, AP-HP and National Academy of Medicine). The author thinks Prof. Nisen Abuaf (Sorbonne University and AP-HP). The author thinks Tech. Said Youssouf Chanfi (Sorbonne University) and Ing. Jean-Michel Bretagne (AP-HP). The author acknowledges Mrs. Norri Zahra and Mr. Regragui Moumaris. The author thinks Clr. Marie-Hélène Maës and SG. Monique Abi (Research and Development Company). The author thinks Danièle Giazzi (Mayor of the 16th arrondissement of Paris 2017-2020).
References
-
Quansah N, Charital S, Yamaryo-Botté Y, Botté CY (2024) Complex Endosymbiosis II: The Nonphotosynthetic Plastid of Apicomplexa Parasites (The Apicoplast) and Its Integrated Metabolism. Methods Mol Biol 2776: 43- 62.
-
Biddau M, Sheiner L (2019) Targeting the apicoplast in malaria. Biochem Soc Trans 47(4): 973-983.
-
Maclean AE, Hayward JA, Huet D, van Dooren GG, Sheiner L (2022) The mystery of massive mitochondrial complexes: the apicomplexan respiratory chain. Trends Parasitol 38(12): 1041-1052.
-
Low LM, Stanisic DI, Good MF (2018) Exploiting the apicoplast: apicoplast-targeting drugs and malaria vaccine development. Microbes Infect 20(9-10): 477- 483.
-
Kennedy K, Crisafulli EM, Ralph SA (2019) Delayed Death by Plastid Inhibition in Apicomplexan Parasites. Trends Parasitol 35(10):747-759.
-
Bulloch MS, Huynh LK, Kennedy K, Ralton JE, McConville MJ, Ralph SA (2024) Apicoplast-derived isoprenoids are essential for biosynthesis of GPI protein anchors, and consequently for egress and invasion in Plasmodium falciparum. PLoS Pathog 20(9): e1012484.
-
Arisue N, Hashimoto T (2015) Phylogeny and evolution of apicoplasts and apicomplexan parasites. Parasitol Int 64(3): 254-259.
-
Shears MJ, Botté CY, McFadden GI (2015) Fatty acid metabolism in the Plasmodium apicoplast: Drugs, doubts and knockouts. Mol Biochem Parasitol 199(1-2): 34-50.
-
Moumaris M (2024) Cell Membrane Compartmentalization and Membrane Dynamics during Plasmodium Infection. Int J Zoo Animal Biol 7(6): 000637.
-
Moumaris M (2024) Advancements in Magnetic Resonance Imaging: Transforming Non-Invasive Diagnosis and Treatment Monitoring in Radiology. J Life Sci Res and Rev 2(5): 1-3.
-
Moumaris M (2024) Plasmodium’s Secret: How a Complex Endomembrane System Drives Malaria’s Deadly Efficiency. Int J Zoo Animal Biol 7(6): 000631.
-
Moumaris M (2024) Malaria’s Hidden Weapon: How Plasmodium Transforms Red Blood Cells to Evade and Invade. Int J Zoo Animal Biol 7(6): 000629.
-
Moumaris M (2024) Advancements in Diagnosing and Treatments Plasmodium knowlesi: Challenges and Innovations. Int J Zoo Animal Biol 7(5): 000622.
-
Moumaris M (2024) Confronting Plasmodium knowlesi: Challenges and Strategies in Malaria Healthcare. Int J Zoo Animal Biol 7(4): 000607.
-
Moumaris M (2024) Unraveling the Enigma: Tackling Knowlesi Malaria in Southeast Asia. Int J Zoo Animal Biol 7(2): 000585.
-
Moumaris M (2024) Unveiling the Enigmatic Plasmodium knowlesi: Insights, Challenges, and Promises in Malaria Research. Int J Zoo Animal Biol 7(1): 000566.
-
Moumaris M (2024) Unlocking the Potential: Overcoming Challenges in CAR-T Cell Therapy for Cancer Treatment. J Biotechnology and Bioprocessing 5(2): 2766-2314.
-
Moumaris M (2023) Revolutionizing Malaria Research: CRISPR unveils New Frontiers. J Biotechnology and Bioprocessing 4(5): 2766-2314.
-
Moumaris M (2024) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [4/4]. Int J Zoo Animal Biol 7(1): 000549.
-
Moumaris M (2023) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [3/4]. Int J Zoo Animal Biol 6(4): 000500.
-
Moumaris M (2023) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [2/4]. Int J Zoo Animal Biol 6(2): 000465.
-
Moumaris M (2022) Lyme Disease: A Zoonosis Tick- Borne Borrelia Bacterium [1/4]. Int J Zoo Animal Biol 5(6): 000425.
-
Moumaris M, Bretagne JM, Abuaf N (2020) Nanomedical Devices and Cancer Theranostics. The Open Nanomedicine and Nanotechnology Journal 6: 1-11.
-
Moumaris M, Bretagne JM, Abuaf N (2019) Biological Membranes and Malaria-Parasites. The Open Parasitology Journal 7: 1-18.
-
Moumaris M, Bretagne JM, Abuaf N (2018) Hospital Engineering of Medical Devices in France. The Open Medical Devices Journal 6: 10-20.
-
Moumaris M, Rajoely B, Abuaf N (2015) Fluorescein Isothiocyanate-Dextran can track Apoptosis and Necrosis induced by heat shock of Peripheral Blood Mononuclear Cells and HeLa Cells. Open Biological Sciences Journal 1: 7-15.
-
Moumaris M, Rajoely B, Abuaf N (2012) The Naïve B Cells are the Lymphocytes with the Highest Anionic Phospholipid Binding Ratios. The Open Immunology Journal 5: 27-35.
-
Moumaris M (2007) Magnetic resonance imaging at the Hôtel-Dieu of Paris. Paris-Descartes University, Unités de formation et de recherche (UFR) biomédicale des Saints- Pères, France.
-
Moumaris M (2005) Identification of a new molecule to monitor apoptosis. Sorbonne-Paris-Nord University, France.
-
Moumaris M (2003) Biomedical research, the law of bioethics relating to the donation and use of elements and products of the human body. Paris-Descartes University, Faculté de Médecine, France.
-
Moumaris M, Abuaf N (2002) Use of labeled dextran for in-vitro assessment of increased cell permeability, cell death and apoptosis. Bulletin officiel de la propriété industrielle (Brevet n°00/09235) 2811682: A3.
-
Moumaris M, Benoliel S, Rouquette AM, Rajoely B, Abuaf N (2000) Phospholipid binding proteins on the plasma membrane of lymphocytes. J Autoimmun 15(2): 81-271.
-
Moumaris M, Ignoti S, Benoliel S, Oghina G, Rajoely B, et al. (1999) Characterization of B-cell adhering to the lamellar phospholipids. French Congress of Antiphospholipid Antibody, Paris, France.
-
Moumaris M (1996) Membranes érythrocytaires dans le paludisme: modèle d’étude: Souris- Plasmodium berghei anka. Université Pierre et Marie Curie, Faculté de Médecine, Paris, France.
-
Moumaris M, Sestier C, Miltgen F, Halbreitch A, Gentilini M, et al. (1995) Effect of Fatty Acid Treatment in Cerebral Malaria-Susceptible and Nonsusceptible Strains of Mice. The Journal of Parasitology 81(6): 997-999.
-
Sabolovic D, Moumaris M, Miltgen F, Sestier C, Halbreich A (1995) A subpopulation of red blood cells induced by bleeding or mosquito sucking. Chinese National Congress of Medical Biophysics, Shanghai, China.
-
Sabolovic D, Moumaris M, Miltgen F, Sestier C, Halbreich A (1995) Characterisation of subpopulation of red blood cells as a preferential target for malaria invasion. French Congress of Electrophoresis, Cell Electrophoresis, Pastor Institute, Paris 19(7): 1215-1219.
-
Moumaris M (1994) Effet des acides gras sur la malaria cérébrale chez des souris susceptible et non susceptible. Université Paris XII, Faculté de Médecine, Créteil, France.
-
Moumaris M (1992) Lyme disease: Serological study. University of Orleans, France.
- Mitochondrial Bio-Logistics: Steering Co-Enzyme Q10 and Lycopene Synergies within the Science 4.0 Bio-OS Framework
- Hymenoptera Specimens from the Caño Negro Wetland, of the National Museum Collection, Costa Rica
- Science 4.0: Comprehensive Architecture of the Biological Operating System (Bio-OS) A Framework for Systemic Resilience and Industrialized Bio-Governance
- Rabbit on, or Hare Back? Understanding Climate Change
- Clinical Validation of Science 4.0: Flow Steering and Epigenetic Drift Inversion on a 76-Year-Old Hybrid System
- Seeds Planted by another Mind