Effects on Cellular Dynamics in the Culture on Human Osteosarcoma Cells of Conditioned Medium Obtained from TGF-B1 Gene Transfected Human Dental Pulp-Derived Mesenchymal Stem Cells
The aim of this study is to suppress the biological properties of tumor cells such as proliferation, apoptosis, cell cycle in osteosarcoma cancer cell line by using the secretome obtained from TGF-B1 transfected DP-MSCs. In addition, another goal of ours is to induce membrane depolarization in mitochondria in osteosarcoma cancer cell line with TGF-B1-CM. In this study, secretomes released from human dental pulp-derived mesenchymal stem cells (hDP-MSCs) with TGF-B1 gene therapy were applied on HOS cell lines. Viability, apoptosis, mitochondrial membrane depolarization and cell cycle analyzes were performed at 24, 48 and 72 hours after the application. Changes in biological characteristics were measured in the Muse Cell Analyzer device according to the product procedure. Statistical analyzes were performed using GraphPad Prism statistical software. TGF-B1 transfected DP-MSC secretome (TGF-B1-CM) reduced the number of viable cells in in vitro cultures of HOS cells. Apoptosis was induced in HOS cells by administration of TGF-B1-CM. Mitochondrial membrane depolarization assays supported the apoptosis results. TGF-B1-CM administration played a role in arresting HOS cells at the G0 / G1 phase of the cell cycle. TGF-B1 transfected DP-MSC secretome suppressed osteosarcoma cells in vitro culture. Considering these results, further molecular studies targeting signaling pathways can be performed.
Hasan Salkin1,2* and Zeynep Burçin Gönen2
Introduction
Osteosarcoma is the most common primary malignant tumor of bone and has an incidence of approximately one in three of a million cases per year worldwide. While it occurs primarily in children and adults, the incidence reaches its second peak above the age of 50 [1]. Osteosarcomas occur at the edges of long bones and metaphyseal growth plates. It is commonly seen in the femur, tibia and humerus, and less commonly in the jaw and pelvis bones. Osteosarcomas are caused by malignancy of osteoblasts that produce immature bone or osteoid. Histologically, it is divided into subtypes such as conventional, low-grade central, periosteal, parasteal, telangiactatic and chondroblastic and small cell form. Some of these histological forms exhibit distinct molecular and biological behavior [2]. Before the 1970s, surgery was generally used in the treatment of osteosarcoma, but with only surgery, the life quality of the patients improved by approximately 15-17% [3, 4]. The transition from surgery to chemotherapy has led to a dramatic improvement in the prognosis of patients with osteosarcoma. Long-term survival rates of less than 20% increased to 65-70% with chemotherapy. However, these rates can be increased when chemotherapy agents (such as methotrexate, doxorubicin and cisplatin) are administered in high doses. Although high doses of chemotherapy agents have a significant effect for treatment, exposure of patients to these agents in high doses also causes other serious pathological consequences and remains a controversial issue today [5]. The introduction of biological agents such as Muramil tripeptide and the use of additional cytotoxic chemotherapy such as ifosfamide did not definitively improve the survival of osteosarcoma patients [5]. Recent advances in molecular genetic studies of osteosarcoma have shown that they change views about the cause of the disease and ongoing therapeutic approaches for patients [6]. In this context, the use of cellular therapy or cellular secretory products instead of chemotherapy agents may be more effective. In recent years, studies have been reported stating that the factors released from mesenchymal stem cells (MSCs) have anti-tumorigenic effects and suggest that these secretion factors can be used as therapeutic agents [7]. TGF-Beta plays a dual role in cancer depending on the stage, and the dual role here indicates the effect that varies from tumor suppressor function to oncogenic function. The mechanisms about these two aspects of TGF-Beta, which cannot be fully elucidated in cancer cells, have caused this situation to be described as “TGF- Beta paradox” by researchers. Clarifying the TGF-Beta paradox will perhaps be important in understanding many of the cancer-related pathways [8, 9, 10, 11]. TGF-B1 is a growth factor that affects important physiological processes such as cell growth, apoptosis, and tissue repair [12]. Instead of applying TGF-B1 as a recombinant protein to osteosarcoma cancer cells, secreting TGF-B1 from MSCs and applying these cells conditional media (TGF- B1-CM) to osteosarcoma cancer cells made us think that it would create more tumor suppressing effects and the study was basically built on this. The aim of this study is to suppress the biological properties of tumor cells such as proliferation, apoptosis, cell cycle in osteosarcoma cancer cell line by using the secretome obtained from TGF-B1 transfected DP-MSCs. In addition, another goal of ours is to induce membrane depolarization in mitochondria in osteosarcoma cancer cell line with TGF- B1-CM. We aimed to develop a therapeutic and biological product that can be used in osteosarcoma cancer treatments and has a better shelf life than cells.
Materials and Methods
Culture of HDP-MSCs
hDPSCs were obtained from Erciyes University Genome and Stem Cell Center, Kayseri, Turkey. The cells were thawed rapidly in a 37-degree water bath, and were transferred into plates at a density of 5,000 cells/cm2 in α-MEM (Biological Industries, Beit-Haemek, Israel) supplemented containing 2 mM L-glutamine (Gibco, USA), 100 U/ml penicillin,100 μg/ml streptomycin (Lonza, MD, USA), 20% Fetal Bovine Serum (Biological Industries, Beit-Haemek, Israel) and 100 μM L-ascorbic acid 2-phosphate (Sigma-Aldric, USA). All the cell cultures were maintained at 37oC in a humidified atmosphere containing 5% CO2. The culture medium was changed every 2∼3 days, and the cell cultures were monitored regularly under an inverted microscope (Leica). When the cells reached 70%∼80% confluence, they were harvested by using 0.05% Trypsin/EDTA (Lonza, MD, USA) and were sub-cultured for further experiments.
Flow Cytometry
The DP-MSCs in the second passage removed from the flask base by trypsinization were resuspended in PBS at a density of 1x106 cells / ml, then the cells were fixed at room temperature for 10 minutes with 10% paraformaldehyde. Cells were blocked with 1% BSA (Bovine serum albumin) for 1 hour. MSC antibodies (CD90-FITC, CD44-PE, CD105 PerCP5.5, CD73-APC and CD34-FITC, CD45-PE, CD11b- PerCP5.5, HLA-DR-APC) (BD Bioscience , Heidelberg, Germany) were prepared in 1% BSA . Measurements were made using a Navios (BeckmanCoulter, USA) flow cytometry device. The data were analyzed with the software KALUZA (BeckmanCoulter, USA).
TGF-B1 Transfection to DP-MSCs
TGF-B1 transfected DP-MSCs, which were produced in our previous study and whose transfection efficiency was determined by us, were used in this study to collect TGF-B1 transfected DP-MSC secretome [13].
The Preperation of TGF-B1 Transfected DP- MSCs-Derived Secretome
In the third passage, TGF-B1 transfected DP-MSCs were grown in standard growth media until they reached 80% confluency. When they reached 80% confluency, standard growth media were removed, serum-free fresh medium was added and cells were conditioned for 18-24 hours in this serum-free medium. At the end of this period, the conditioned media from TGF-B1 transfected DP-MSCs were collected in 50 ml polypropylene tubes. It was centrifuged at 3000 x g for 25 min at + 4oC in a refrigerated centrifuge to remove dead cells and cellular debris from the environment. The supernatant was then transferred to clean tubes. Then, the obtained conditional media were filtered with 0.22 µm sterile filters.
Culture of HOS Cell Lines
The HOS cell line was supplied cryopreserved by purchase as a ready-made cell line from ATCC (ATCC® CRL-1543) and cultured according to the product protocol.
Therefore, an ethics committee document was not needed in our study. Cells were quickly thawed in a 37oC water bath and supplemented with 2 mM L-glutamine (Gibco, USA) and 100 U / ml penicillin, 100 μg / ml streptomycin (Lonza, MD, USA) and 10% Fetal Bovine Serum (FBS). It was added to flasks at a density of 5000 cells / cm2 in Eagle’s Minimum Essential Medium (EMEM). Cells were allowed to grow in an incubator at 37oC and humid atmosphere containing 5% CO2. The old medium was replaced with fresh medium every 3 days. When the proliferated cells reached 70% confluency, they were removed from the environment by trypsin using 0.05% Trypsin / EDTA. Thus, subcultures were made up to the number of cells required for the tests we will perform in further experiments.
Application of Conditional Media on HOS Cell Lines
Eagle’s Minimum Essential Medium (EMEM), 10% FBS, 1% Penicillin-streptomycin and 2mM glutamax were used as standard growth culture medium for HOS cell lines. Then, conditional media obtained from TGFB1 transfected DP- MSCs (TGF-B1-CM) were applied on HOS cell lines for 24, 48 and 72 hours.
Analysis of Viability and Cell Count
Viability percentages of cells were determined with the Muse Cell Analyzer (Merck Millipore) cell count and viability kit (Muse Count & Viability Kit MCH100102).
Annexin V Assay
HOS cells were inoculated into 6-well culture dishes at 5000 cells / cm2 in standard growth media. After 24 hours, TGF-B1-CM was applied by removing the media from the cells. In order to determine the percentage of apoptosis in cells at 24, 48 and 72 hours after the application, annexin V, a marker that binds to phosphatidylserine released in membrane structures in cells undergoing apoptosis, was used. Total apoptosis percentages in the cells were measured according to the product procedure in the Muse Cell Analyzer (Millipore) device operating with flow cytometry logic using annexin V and cell death kit (Annexin V and Dead cell kit, Muse, Millipore). All experiments were run in triplicate.
Determination of Mitochondrial Membrane Depolarization
A cationic and lipophilic dye was used to determine the membrane potential of the mitochondrial membrane in HOS cells at 24, 48 and 72 hours. Mitochondrial membrane depolarization was measured in the Muse Cell Analyzer (Millipore) device working with flow cytometry principle using the mitochondrial potential test kit (Muse Mitopotential Assay kit, MERCK, Millipore). Cells were collected in microcentrifuge tubes at 1 x 105 cells. It was diluted in a volume of 100 µl with 1 x test buffer in the kit and 95 µl cationic-lipophilic dye solution was added. After vortexing for a short time, it was incubated for 20 minutes at 37oC in an incubator containing 5% CO2. After the incubation, 7-AAD reagent was added to each tube to mark dead cells. Then it was incubated at room temperature for 5 minutes. Samples prepared according to the product protocol were measured. All experiments were run in triplicate.
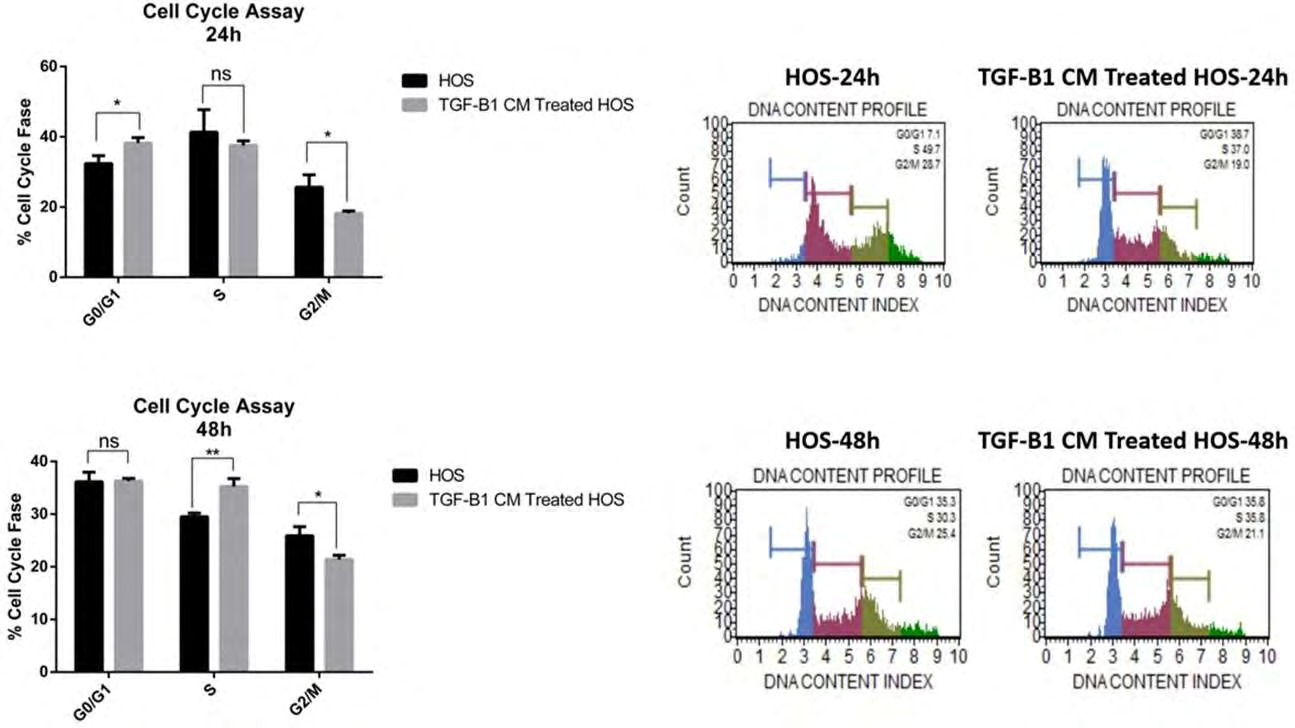
Cell Cycle Assay
HOS cells were inoculated into 6-well culture dishes at 5000 cells / cm2 in standard growth media. After 24 hours, TGF-B1-CM was applied by removing the media from the cells. Cells in each group were collected separately in microcentrifuge tubes at 24 and 48 hours after the application. After washing twice with PBS, the PBS was removed by centrifugation at 350 x g for 5 minutes and fixed with 70% ethanol for 3 hours at -20°C. After fixation, it was washed again with PBS and cells were treated in hypotonic buffer solution containing propodium iodide. Then, cell numbers and ratios in G1 / G0, S and G2 / M phases were measured according to the product procedure by using the cell cycle test kit (Cell Cycle Assay kit, Muse, Millipore) using the Muse Cell Analyzer (Millipore) device working with flow cytometry principle. All experiments were run in triplicate.
Statistical Analysis
Biological activity analyzes among the groups were analyzed by un-paired t-tests. Multiple comparison analyzes between groups for each test were determined by Multiple t-tests. Statistical analysis was performed using the “Graph Pad Prism Version 6” statistical software. Data with P <0.05 were considered statistically significant.
Results
Culture and Characterization of DP-MSCs
It was determined that DP-MSCs showed typical fibroblastic and star-shaped morphology in culture. Flow cytometry analysis showed that DP-MSCs positively expressed MSC markers. DP-MSCs were negative in terms of hematopoietic stem cell markers (Figure 1).
TGF-B1 Gene Transfection
TGF-B1 Transfected DP-MSCs (13), which were successfully produced in our previous study and had 95% transfection efficiency, were used in this study (Figure 1).
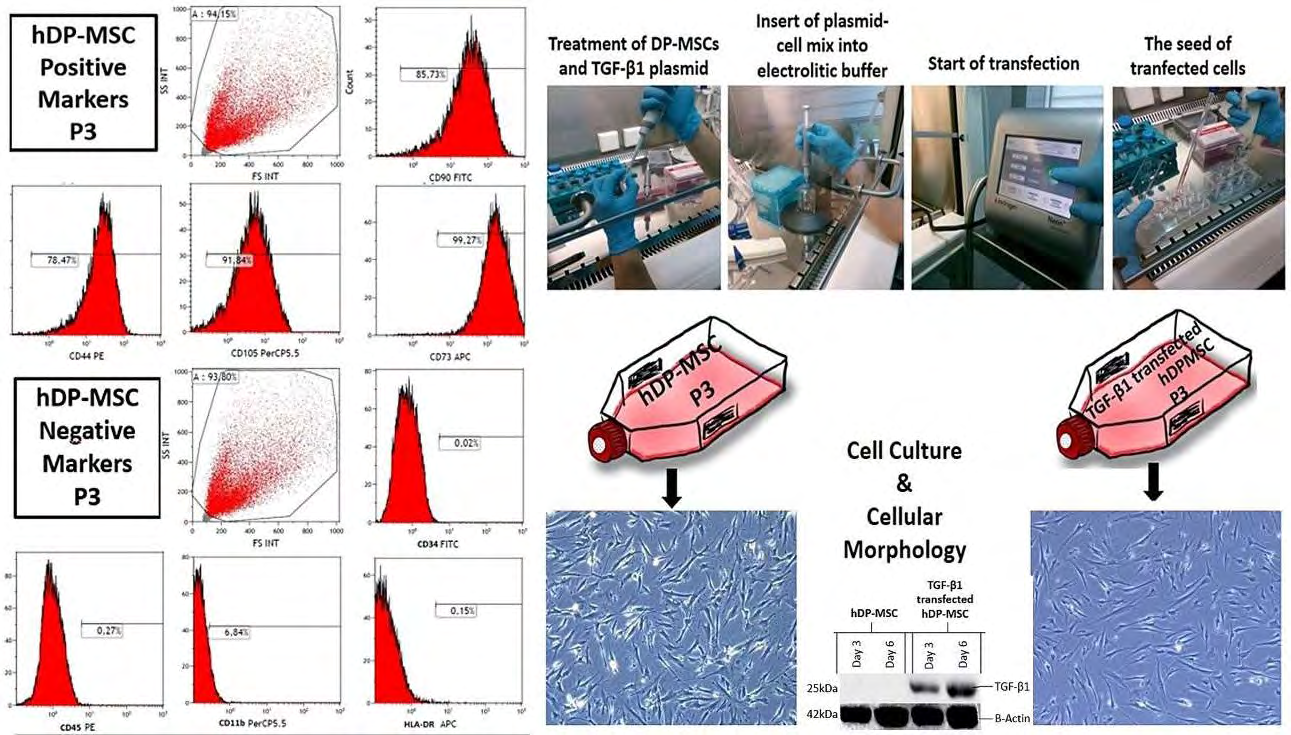
Collection of TGF-B1 Transfected DP-MSCs- derived Secretome
Sufficient amount of conditional media was successfully collected from TGF-B1 transfected DP-MSCs. Conditioned media were applied freshly to HOS cell lines. Secretome applications to HOS cell lines were successfully performed in standard growth media when they reached 80% confluency.
Analysis of Viability and Cell Count
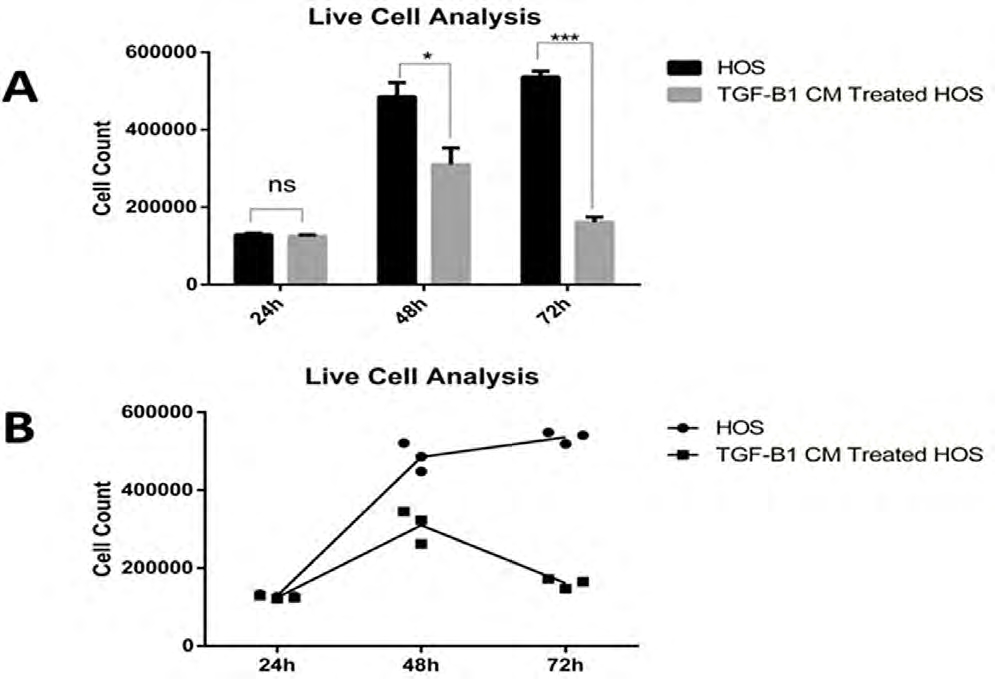
Viability assays significantly decreased cell numbers in HOS cell lines treated with TGF- B1-CM, especially at 48 and 72 hours. The results were statistically significant (p <0.05). There was no statistically significant difference when compared with the control group at 24 hours (Figure 2).
Annexin V Assay
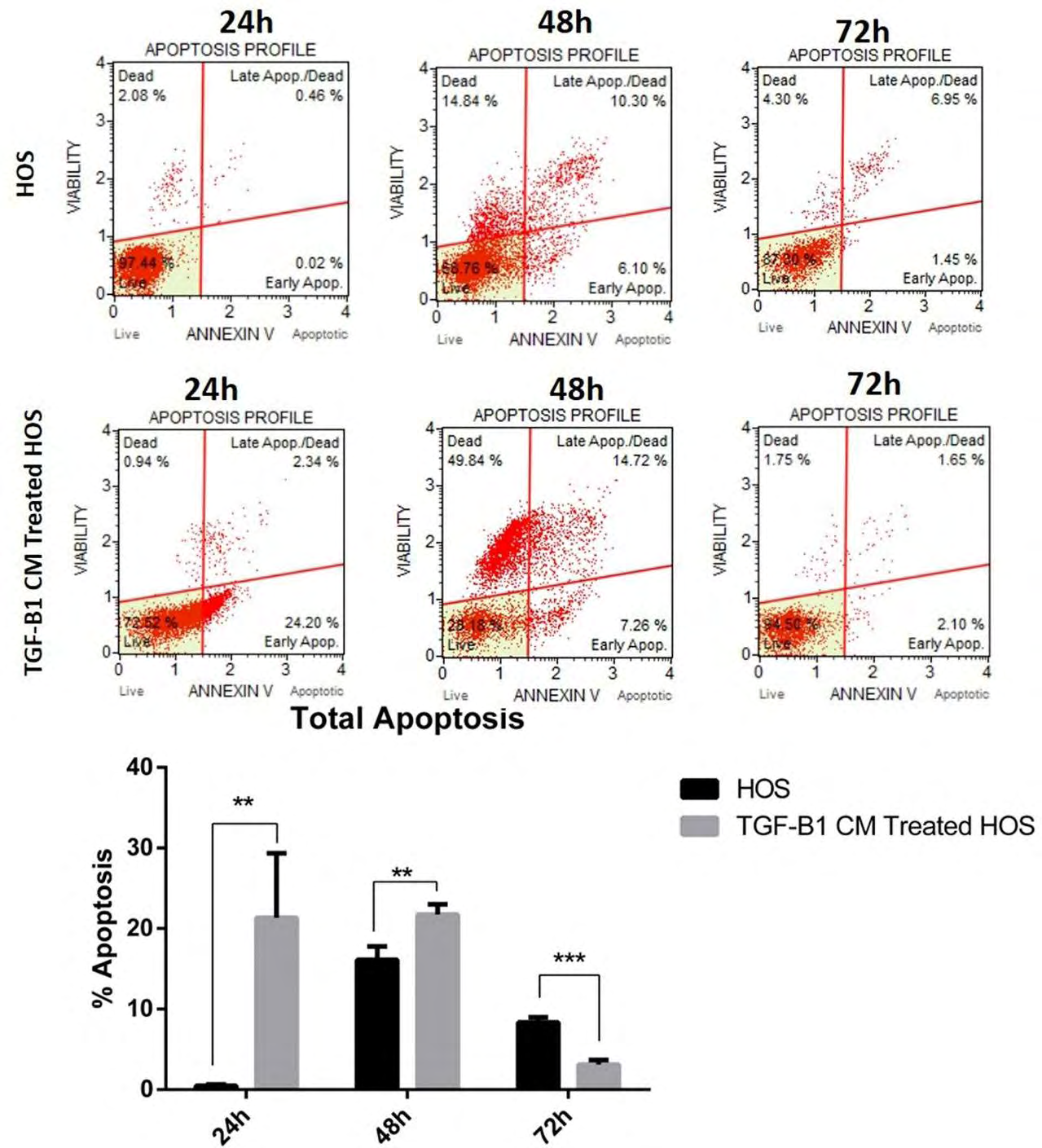
While total apoptosis was 0.495% in the control group at the 24th hour, it was measured as 21.345% in the TGF- B1-CM group (p = 0.00201849). After the 48th hour analysis, the mean total apoptosis was 16.1% in the control group and 21.75% in the TGF-B1-CM group (p = 0.00940457). Apoptosis at 72 hours was 8.35% in the control group and 3.13% in the TGF-B1-CM group (p = 0.00039). At 24 and 48 hours, apoptosis was significantly induced in HOS cells by TGF-B1 transfected DP-MSC-derived secretome (Figure 3).
Mitochondrial Membrane Depolarization
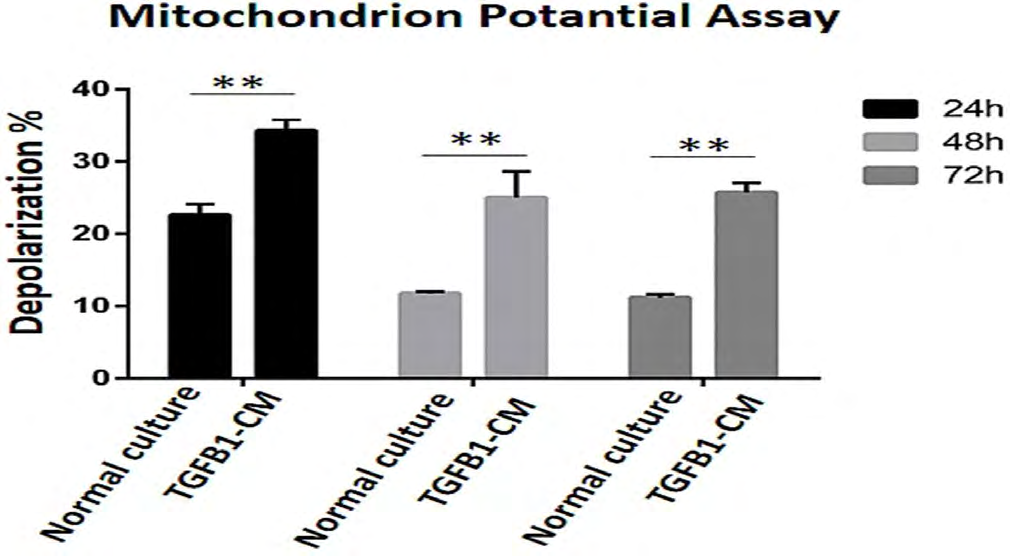
TGF-B1-CM application on HOS cell lines significantly increased mitochondrial membrane depolarization at 24, 48 and 72 hours (Figure 4).
Cell Cycle Assay
In HOS cell lines, TGF-B1-CM application caused the arrest of HOS cells in the G0 / G1 phase at 24 hours. Both the number of cells in the S stage and the percentage of cells in the M stage decreased with the application of TGF-B1-CM. The decrease in the percentage of cells in the M phase of the cell cycle continued in the 48th hour analysis (Figure 5).
Discussion
Acquiring important biological functions by releasing the TGF-B signaling pathway by cancer cells has recently emerged as a leading area of preclinical and clinical cancer research. In early stage tumors, the TGF-B pathway promotes cell cycle arrest and apoptosis. In contrast, in advanced stages, it promotes tumor progression and metastasis by promoting cancer cell motility, invasion, epithelial- mesenchymal transition (EMT), and cell resilience [14, 15]. This contradictory dual role is known as the “TGF-B paradox” [16]. Osteosarcomas are not only diverse in terms of their pathological nature and clinical distribution but also in the aspect of the cellular heterogeneity within the tumors. Osteosarcoma most commonly develops around the knee joint, distal femur, and proximal tibia; the vast majority of the patients have already shown micrometastasis at the time of diagnosis [17]. It is believed to originate from bone mesenchymal cells or osteoprogenitors [18]. The etiology of osteosarcoma is currently rather limited, while the pathogenesis remains unknown. The unique properties of osteosarcoma may be related to either the cell of origin or components in the bone marrow microenvironment, such as the large amount of transforming growth factor b1 and low oxygen tension [19, 20]. Once an osteosarcoma germinates in the bone, tumor cells secrete factors that initiate osteoclast- mediated bone destruction, and matrix-derived growth factors, especially TGF-B1, are released from bone matrix. At this time, osteosarcoma cells also release TGF-B1 directly [21, 22]. The biological action of the TGF-Beta signaling pathway is mediated by type I and type II receptors (TGFBRI and TGFBRII) [23]. The canonical pathway involves the activation of SMAD paths [24]. TGF-B binds first to TGFBRII, which targets and activates TGFBRI [23, 24, 25]. It then activates Smad2 / 3 by phosphorylating. Activated SMAD2 / 3 combines with SMAD4 and migrates to the nucleus to regulate transcription [26]. In addition to the SMAD pathway, TGF-B also signals in a number of non-canonical pathways, including m-TOR, RhoA, Ras, MAPK, PI3K / AKT, PP2A / p70s6K, and JNK [27]. The relative importance and interaction of these TGF-B signaling pathways are still under investigation [28, 29]. Therefore, secretomas of mesenchymal stem cells derived from TGF-B1 transfected dental pulp were used in our study. The fact that TGF-B1 is one of the apoptosis regulators and its relationships with many other cellular pathways have made us think that HOS cells can be further suppressed in in vitro cultures in this way. Our apoptosis and cell cycle analyzes have supported this. In addition, it was found in our study that apoptosis was induced in accordance with the time periods in which mitochondrial membrane depolarization was induced in HOS cells. Zamilpa et al showed that especially c-terminal fragments of TGF-B1 protein are required for apoptosis in human osteosarcoma cells. They reported that this interaction occurred through TGF-B1 protein c-terminal fragments and integrins and initiated apoptosis induction [30]. The use of stem cell secretomes containing TGF-B1 ligands in our study supported this interaction, but unlike this study, TGF-B1 ligands were applied to HOS cells together with the stem cell secretome, and cancer cells were more actively suppression provided with both secreted TGF-B1 and MSC secretion factors. In 2000, the concept of “Dental Pulp Stem Cells” was added to the literature by Gronthos et al. with pulpal stem cells obtained from human 3rd molar teeth. It has become very popular as a promising, potential cell in terms of its rich biologic features, which have been studied on many occasions as of that date [31]. Dental Pulp Stem Cells can be used in both in vitro and in vivo clinical, surgical and tissue engineering studies. In addition, it is a very important advantage for DP-MSCs that plasmid DNA can be transferred easily in culture and that desired cytokines such as TGF-B1 can be obtained from cultures easily. Therefore, DP-MSCs were preferred as stem cell source in our study. It is now known that cell interactions through secretomes, rather than direct interaction with the stem cell used, create more specific effects in terms of both regenerative and cancer medicine, and the new trend has been stem cell secretomas. Secretomes released from MSCs are a very rich secretion product in terms of growth factors and TGF-B1 content [32, 33]. The suppressive effect of MSC conditioned environments on cancer cells makes us think that it can be used more effectively with some genetic modifications in anti-tumor studies. Therefore, the combination of TGF-B1 overexpressing MSC secretome may be much more effective in suppressing cancer cells instead of using only TGF-B1 recombinant protein or MSC conditional medium alone. Therefore, in our study, we aimed to obtain a secretome containing more TGF-B1 protein by transferring the TGF- B1 gene through gene transfer to DP-MSCs. When we applied the secretomes we obtained to HOS cultures, we found that we suppressed the biological characteristics in osteosarcoma cancer cells. TGF-B1 transfected DP-MSC secretome (TGF-B1-CM) reduced the number of viable cells in in vitro cultures of HOS cells. Apoptosis was induced in HOS cells by administration of TGF-B1-CM. Mitochondrial membrane depolarization assays supported the apoptosis results. TGF-B1-CM administration played a role in arresting HOS cells at the G0 / G1 phase of the cell cycle. As a result, TGF-B1 transfected DP-MSC secretome suppressed osteosarcoma cells in vitro culture. Considering these results, further molecular studies targeting signaling pathways can be performed.
Conclusion
Our results showed that the culture and characterization of DP-MSCs were successfully performed. The TGF-B1 gene transferred DP-MSCs produced in our previous study were used in this study (13). TGF-B1 transfected DP-MSC secretomes were collected from these transfected cells in a standard manner and successfully applied to HOS cell lines. Viability in HOS cell lines was suppressed at 48 and 72 hours after the applications. Apoptosis increased significantly in HOS cell cultures at 24 and 48 hours. The results of apoptosis supported the results of mitochondrial membrane depolarization. In HOS cell cultures with DPSC secretome, arrest occurred in the cell cycle, especially at the G0 stage. Considering all these results, it has been seen that TGF-B1 transfected DPSC secretomes produce suppressive effects in HOS cell lines. Further studies can be done specifically targeting signal pathways.
Conflict of Interest
The authors have declared that no conflict of interest exists.
Acknowledgements
This study was performed in Erciyes University Genome and Stem Cell Center, Kayseri- Turkey. Authors thank to this institution for its support.
References
-
WHO (2013) WHO Classification of Tumours of Soft Tissue and Bone, 4th (Edn.), International Agency for Research on Cancer, pp: 281-295.
-
Kansara M, Teng MW, Smyth MJ, Thomas DM (2014) Translational biology of osteosarcoma. Nat Rev Cancer 14(11): 722-735.
-
Bernthal NM, Federman N, Eilber FR, Nelson SD, Eckardt JJ, et al. (2012) Long-term results (>25 years) of a randomized, prospective clinical trial evaluating chemotherapy in patients with high-grade, operable osteosarcoma. Cancer 118(23): 5888-5893.
-
Link MP, Goorin AM, Miser AW, Green AA, Pratt CB, et al. (1986) The effect of adjuvant chemotherapy on relapse- free survival in patients with osteosarcoma of the extremity. N Engl J Med 314(25): 1600-1606.
-
Isakoff MS, Bielack SS, Meltzer P, Gorlick R (2015) Osteosarcoma: Current Treatment and a Collaborative Pathway to Success. J Clin Oncol 33(27):3029-3035.
-
Rickel K, Fang F, Tao J (2017) Molecular genetics of osteosarcoma. Bone 102: 69-79.
-
Cheng L, Zhang K, Wu S, Cui M, Xu T, et al. (2017) Focus on Mesenchymal Stem Cell-Derived Exosomes: Opportunities and Challenges in Cell-Free Therapy. Stem Cells Int 2017: 6305295.
-
Humbert L, Neel JC, Lebrun JJ (2010) Targetting TGF-B signaling in human cancer therapy. Trends in Cell Mol Biol 5: 69-107.
-
Dai M, Al-Odaini AA, Arakelian A, Rabbani SA, Ali S, et al. (2012) A novel function for p21Cip1 and acetyltransferase p/CAF as critical transcriptional regulators of TGFβ- mediated breast cancer cell migration and invasion. Breast Cancer Res 14(5): R127.
-
Dai M, Al-Odaini AA, Flis-Aime N, Villatoro MA, Guo J, et al. (2013) Cyclin D1 cooperates with p21 to regulate TGFβ-mediated breast cancer cell migration and tumor local invasion. Breast Cancer Res 15(3): R49.
-
Neel JC, Humbert L, Lebrun JJ (2012) The Dual Role of TGF𝛽 in Human Cancer: From Tumor Suppression to Cancer Metastasis. ISRN Mol Biol 2012: 1-28.
-
Wakefield LM, Hill CS (2013) Beyond TGFbeta: roles of other TGFbeta superfamily members in cancer. Nat Rev Cancer 13(15): 328-341.
-
Salkın H, Gönen ZB, Ergen E, Bahar D, Çetin M, et al. (2019) Effects of TGF-β1 Overexpression on Biological Characteristics of Human Dental Pulp-derived Mesenchymal Stromal Cells. Int J Stem Cells 12(1): 170- 182.
-
Jakowlew SB (2006) Transforming growth factor-beta in cancer and metastasis. Cancer Metastasis Rev 25(23): 435-457.
-
Drabsch Y, Ten Dijke P (2012) TGF-beta signalling and its role in cancer progression and metastasis. Cancer Metastasis Rev 31(3-4): 553-568.
-
Wendt MK, Tian M, Schiemann WP (2012) Deconstructing the mechanisms and consequences of TGF-beta-induced EMT during cancer progression. Cell Tissue Res 347(1): 85-101.
-
Mirabello L, Troisi RJ, Savage SA (2009) International osteosarcoma incidence patterns in children and adolescents, middle ages and elderly persons. Int J Cancer 125(1): 229- 234.
-
Mohseny AB, Szuhai K, Romeo S, Emilie P Buddingh, Inge Briaire-de Bruijn, et al. (2009) Osteosarcoma originates from mesenchymal stem cells in consequence of aneuploidization and genomic loss of Cdkn2. J Pathol 219(3): 294-305.
-
Robey PG, Young MF, Flanders KC, Roche NS, Kondaiah P, et al. (1987) Osteoblasts synthesize and respond to transforming growth factor-type beta (TGF-beta) in vitro. J Cell Biol 105(1): 457-463.
-
Rankin EB, Giaccia AJ, Schipani E (2011) A central role for hypoxic signaling in cartilage, bone, and hematopoiesis. Curr Osteoporos Rep 9(2): 46-52.
-
Mundy GR (1997) Malignancy and the skeleton. Horm Metab Res 29(3): 120-127.
-
Zhang H, Wu H, Zheng J, Yu P, Xu L, et al. (2013) Transforming Growth Factor b1 Signal is Crucial for Dedifferentiation of Cancer Cells to Cancer Stem Cells in Osteosarcoma. Stem Cells 31(3): 433-446.
-
Massagué J, Gomis RR (2006) The logic of TGF-β signaling. FEBS Lett 580(12): 2811-2820.
-
Shi Y, Massagué J (2003) Mechanisms of TGF-β signaling from cell membrane to the nucleus. Cell 113(6): 685- 700.
-
Derynck R, Akhurst RJ, Balmain A (2001) TGF-beta signaling in tumor suppression and cancer progression. Nat Genet 29(2): 117-129.
-
Vogelmann R, Nguyen-Tat MD, Giehl K, Adler G, Wedlich D, et al. (2005) TGFβ induced downregulation of E-cadherin-based cell-cell adhesion depends on PI3- kinase and PTEN. J Cell Sci 118(pt20): 4901-4912.
-
Mu Y, Gudey SK, Landström M (2012) Non-Smad signaling pathways. Cell Tissue Res 347(1): 11-20.
-
Miyazono K (2009) Transforming growth factor-beta signaling in epithelialmesenchymal transition and progression of cancer. Proc Jpn Acad Ser B Phys Biol Sci 85(8): 314-323.
-
Gomes LR, Terra LF, Wailemann RA, Labriola L, Sogayar MC, et al. (2012) TGF-β1 modulates the homeostasis between MMPs and MMP inhibitors through p38 MAPK and ERK1/2 in highly invasive breast cancer cells. BMC Cancer 19(12): 26.
-
Zamilpa R, Rupaimoole R, Phelix CF, Somaraki-Cormier M, Haskins W, et al. (2009) C-Terminal Fragement of Transforming Growth Factor Beta-Induced Protein (TGFB1p) Is Required For Apoptosis In Human Osteosarcoma Cells. Matrix Biol 28(6): 347-353.
-
Gronthos S, Mankani M, Brahim J, Robey PG, Shi S, et al. (2000) Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc Natl Acad Sci USA 97(25): 13625-13630.
-
Wu YC, Wang WT, Huang LJ, Cheng RY, Kuo YR, et al. (2019) Differential Response of Non-cancerous and Malignant Breast Cancer Cells to Conditioned Medium of Adipose tissue-derived Stromal Cells (ASCs). Int J Med Sci 16(6): 893-901.
-
Jung H, Kim HH, Lee DH, Shik Y, Yang HC, et al. (2011) Transforming growth factor- beta 1 in adipose derived stem cells conditioned medium is a dominant paracrine mediator determines hyaluronic acid and collagen expression profile. Cytotechnology 63(1): 57-66.
- Research Progress of Induced Pluripotent Stem Cells and Their Clinical Application Prospects
- Nishan Al-Kamal is the Starting Point of A Feminist Scientist
- Current Concepts and Future Perspectives of Stem Cell Therapy in Peripheral Arterial Disease
- Stem Cell and Oxidative Stress-Inflammation Cycle
- Adipose Derived Mesenchymal Stem Cells Origin, Characteristics and Promises
- Mitochondria Targeted Antioxidants can Improve In Vitro Embryo Production in Buffalo