Purification and Biochemical Characterization of a Lipase Produced by Aspergillus Niger: Interfacial Binding, Activity of Lipase at the Lipid-Water Interface Using Monomolecular Film Techniques
Aspergillus Niger lipase (ANL) was purified from a culture medium. Pure ANL was obtained after ammonium sulfate fractionation, Sephacryl S-200 gel filtration and cation exchange chromatography (Mono-Q). The pure lipase, which is not a glycoprotein, was presented as a monomer having a molecular mass of about 99 kDa. The lipase activity was maximal at pH 8 and at 39°C. ANL hydrolyses the long chains of triacylglycerols more efficiently than the short ones. A specific activity of 3779 U/mg was measured on olive oil as substrate at 39°C and at pH 8 in the presence of 4 mM NaTDC. ANL was inactivated when the enzyme was incubated at 75°C or at pH less than 4. Natural detergent (NaTDC), pH dependence of the catalytic activity of ANL was at pH 8, which confirm that ANL present one pH profile.
Faouzi Laachari* and Ibnsouda SK
Université Sidi Mohamed Ben Abdellah, Morocco
Keywords: Aspergillus Niger; Lipase; Purification; pH dependence
Introduction
The enzymatic synthesis of esters, involving in particular lipases, is an interesting alternative that allows to work under milder operating conditions and to have a greater selectivity which limits the formation of byproducts. The lipases, in particular, have been the subject of several studies in which the reactions of esterification, trans esterification and interesterification were studied [1, 2, 3, 4, 5, 6]. In a catalytic process, the stability and the activity of biocatalyst are considered the key parameters affecting the viability of the system. In order to reduce the costs associated with the use of these enzymes. They must be active for long periods, therefore the factors influencing their activity and stability must be well controlled. The activity and stability of enzymes are intrinsically related to their nature, but are also influenced by external parameters such as temperature rise or pressure and extreme micro-environmental pH or poorly controlled water activity [7]. It is interesting to note that some enzymes better retain their catalytic activity at high temperature in a non-aqueous medium and in the presence of hydrophobic solvents in the presence of water [8, 9]. This is due to the fact that in an organic solvent, the enzyme is trapped in its active conformation since water is almost absent which gives the enzyme greater conformational flexibility [9]. The water activity is one of the most important factors that influence the enzyme activity. Indeed, to maintain its activity, an enzyme requires the presence of a specific amount of water in its microenvironment. In fact, water is involved mainly in the maintenance of the active conformation of the enzyme via non-covalent bonds and hydrogen bonds. This therefore relates to the water directly related to the enzyme [8, 9].
Material and methods
Enzymes and Proteins Purification
Aspergillus niger lipase (ANL) was purified in (Laboratoire de Biochimie et de Génie Enzymatique des Lipases, ENIS, BPW 3038 Sfax, Tunisia) [10].
Culture Conditions
The culture medium was precultured during 12 h at 30°C and 150 rpm in 250 ml shaking flasks containing 50ml of medium A (17 g/l casein peptone, 5 g/l yeast extract (Difco), 2.5 g/l glucose, 0,5 g/l MgSO4, 2 g/l CaCl2, 1,5 g/l sodium citrate, 2 g/l KHPO4, pH (5.5). Aspergillus niger cultures used as inocula (2%) were cultivated in 1-l shaking flasks containing 250 ml of medium A. The culture was incubated aerobically during 72 h on a rotary shaker set at 150 rpm and at a temperature of 30°C.
Lipase Activity Determination
The lipase activity was measured titrimetrically at pH 8.5 and at 40°C with a pH-stat under standard conditions using tributyrin (0.25 ml in 30 ml of 2.5 mM Tris-HCl pH 8.2, 2 mM CaCl2, 2 mM NaDC) or olive oil emulsion as substrate (10 ml in 20 ml of 2.5 mM Tris-HCl pH 8.2, 2 mM CaCl2, 4 mM NaDC) [11]. Lipolytic activity was expressed as units. One unit corresponds to 1 µmol of fatty acid released per minute.
Determination of Protein Concentration
Protein concentration was determined as described by Bradford MMA [12] using BSA as standard.
Procedure of ANL Purification
ANL was purified to homogeneity from a culture medium after ammonium sulfate precipitation, Sephacryl S-200 gel filtration and cation exchange chromatographie (Mono Q Sepharose). Pure ANL was eluted as a tetrameric protein (99 kDa) [13].
Monomolecular Film Techniques
Measurements were performed with KSV-2000 Baro-stat equipment (KSV-Helsinki). The principle of the method was described previously Verger R and De Haas GH [14]. It involves the use of a “zeroorder” trough with two compartments: a reaction compartment and a reservoir compartment connected to each other by a small surface channel. The enzyme solution was injected only, into the subphase of the reaction compartment when the lipid film covered both of them. A mobile barrier, automatically driven by the Barostat, moved back and forth over the reservoir to keep the surface pressure (π) constant, thus compensating for the substrate molecules that were removed from the film by enzyme hydrolysis. The surface pressure was measured on the reservoir compartment with a Wilhelmy plate (perimeter 3.94 cm) attached to an electro microbalance, which was connected in turn to a microprocessor programmed to regulate the mobile-barrier movement. As shown previously, the sensitivity of the Wilhelmy plate was estimated to 0.2 mNm-1. The monomolecular film technique was used to measure the difference of the surface tension existing between the surface of an aqueous phase (γ0) and that of the same liquid covered by an amphipatic molecular film (γ). This difference corresponds to the surface pressure (π) [40], π = γ0-γ, where γ0 is the surface tension of aqueous phase (72.8 mNm-1 at 20°C for water) and γ is the surface tension of the same aqueous phase covered by a molecular amphipatic film. Two 2-cm magnetic stirrers stirred the reaction compartment at 250 rpm. The reactions were performed at room temperature (25°C). The surface of the reaction compartment was 108.58 cm2and its volume was 130 ml. The reservoir compartment was 148mm wide and 249mm long. Before each experiment, the Teflon trough used for forming the monomolecular film was cleaned with water, then gently brushed in the presence of distilled ethanol, washed again with tap water and finally rinsed with double distilled water. The aqueous subphase was composed of various buffer prepared with double-distilled water and filtered through a 0.45μm Millipore filter. Any residual surface-active impurities were removed before each assay by a sweeping and suction of the surface. The kinetic data were analyzed as described previously. All the activities measurement was performed after 10 min of lipase injection under different substrate tested. Activities are expressed as the number of moles of substrate hydrolyzed per unit time and unit surface of the reaction compartment of the “zero- order” trough for an arbitrary lipase concentration of 1M.
Measurement of the pH Dependency
The variation with pH in the ANL were measured using rac-dicaprin monomolecular films that were maintained at a constant surface pressure of 35 mN.m-1. Assays were carried out at room temperature in a zero-order. The aqueous subphase was composed of various buffers that contained 150mM NaCl and 21mM CaCl2, 10mM glycine (pH 3-4), 10mM sodium acetate (pH 5-6), 10 mM phosphate (pH 7), 10 mM Tris-HCl (pH 8-9).
Analytical Methods
Analytical polyacrylamide gel electrophoresis of proteins in the presence of sodium dodecyl sulfate (0.3 M) and β-mercaptoethanol (0.25 M) or DTT (0.5 M) (SDS/PAGE) was performed by the method of Laemmli.
Effect of Free Fatty Acids
The lipase activity was measured as a function of various substrate (TC4, olive oil) concentrations ranging from 0-40mM. The Michaelis-Menten constant (Km) and the maximum velocity (Vmax) for the reaction with TC4, TC8 or TC18 as substrate were calculated by Lineweaver-Burk plot.
Effect of Detergents
The lipase activity was measured using tributyrin and olive oil as a substrate in the presence of increasing concentrations of NaDC ranging from 0 to 10 mM, under optimum conditions of pH and temperature.
Effect of pH and Temperature on ANL Activity and Stability
ANL activity was tested in various buffers at different pH (5-10.5) at 40°C. Furthermore, the pH stability of the lipase was measured by incubating the enzyme at different pH (3-12) for 24 h at room temperature. The residual activity was determined, after centrifugation, under standard assay method. Moreover, the optimum temperature for the ANL activity was determined by carrying out the enzyme assay at different temperatures (25-60°C) at pH 8.5. Finally, the effect of temperature on lipase stability was determined by incubating the enzyme solution at different temperatures (30- 80°C) for 30 min and the residual activity was determined, after centrifugation, under standard assay method.
Kinetic Study
Lipase activities were measured by using a various substrate (TC4, TC8 or TC18) concentrations (0-40 mM). The Michaelis–Menten constant (KMapp.) and the maximum velocity (Vmax) were calculated by Lineweaver-Burk plot.
Results and Discussion
Lipase Production
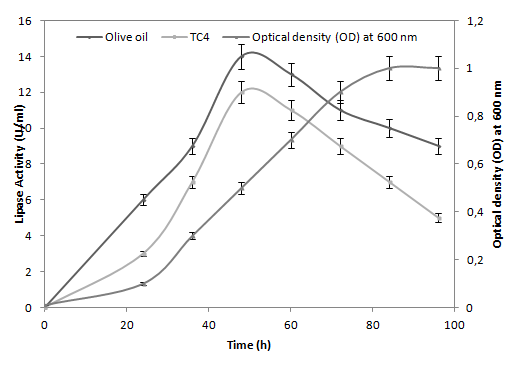
The ANL maximum activity (14 U/ml) was obtained after 48 h incubation (Figure 1). The lipase activity was measured titrimetrically at pH 8 and 39°C with a pH-stat and the time course of lipase production was followed at 30°C with cell growth (Figure 1). Moreover, ANL production was not induced by the presence of triacylglycérols (TC4 or olive oil) or esters (Tween-20) (data not shown).
Purification of ANL The purified lipase was homogenous when tested by the coomassie blue staining in SDS-PAGE (Figure 2). This figure shows that one band was revealed for ANL with a molecular mass of 99 kDa. The purification flow sheet is shown in Table 1. The specific activity of the pure lipase reached 3779 U/ mg when using olive oil emulsified with gum Arabic as substrate at pH 8 and 39°C. Under the same conditions, a specific activity of 3779 U/mg was obtained when using TC4 as substrate.
| Purification Step | Total Activity (U/mL)a | Proteins (mg)b | Specefic Activity (U/ mg) | Activity Recovery (%) | Purification Factor |
|---|---|---|---|---|---|
| Culture Supernantant | 14000±330 | 1400±0,5 | 10 | 100 | 1 |
| (NH ) SO precipitation 4 2 4 | 11900±170 | 910±0,75 | 13±0,35 | 76±0,7 | 1,4±0,03 |
| Sephacryl S 200 | 10170±90 | 60±0,5 | 169±0,25 | 56±0,6 | 12±0,4 |
| Mono-S chromatiography | 7975±55 | 2,11±0,7 | 3779±0,33 | 37±1,9 | 430±1,55 |
Table 1: ANL Flow sheet purification.
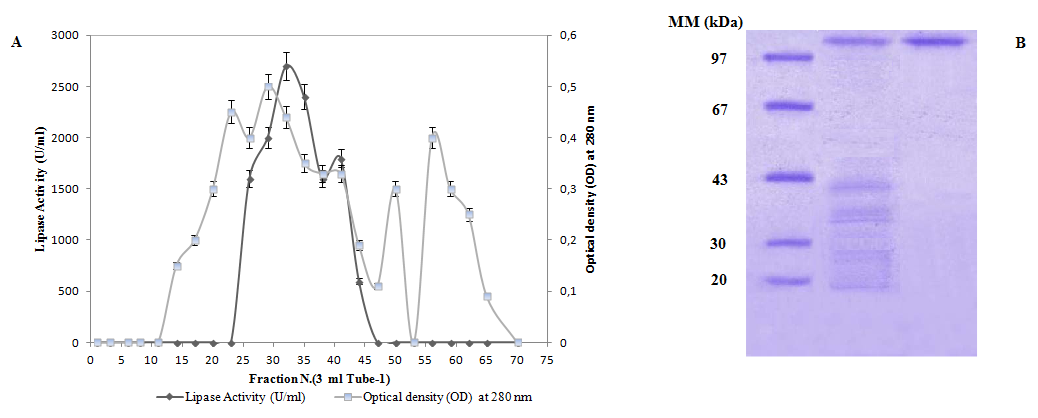
Figure 2: (A) Chromatography of ANL on Sephacryl S-200 gel filtration and cation exchange chromatography (Mono-Q). The colomn (3 × 100 cm) was equilibrated with buffer a (20 mM sodium acetate pH 5.4, 20 Mm NaCl, 1 mM benzamidine). The elution of lipase was performed with the same buffer at a rate of 30 ml/h. Lipolytic activity was measured under standard conditions at pH 8.00 and 50°C using a pH-stat. (B) SDS/PAGE (15%). Lane 1, molecular mass markers (Pharmacia); lane 2, characterisation of the ANL obtained after Sephacryl S-200 chromatography and (Mono-Q).
pH Dependence of the Catalytic Activity of ANL
We tested the pH influence of the aqueous subphase on the ANL activity (Figure 3) using rac-dicaprin monomolecular films that were maintained at a constant surface pressure of 35 mN.m-1. From the figure 3, we noted that the maximal activity of ANL is reached at pH 8, similar results were obtained using olive oil emulsion as a substrate with the pH- stat assay. Comparable results were obtained by using ANL [15]. These results confirm that ANL present one pH profile.
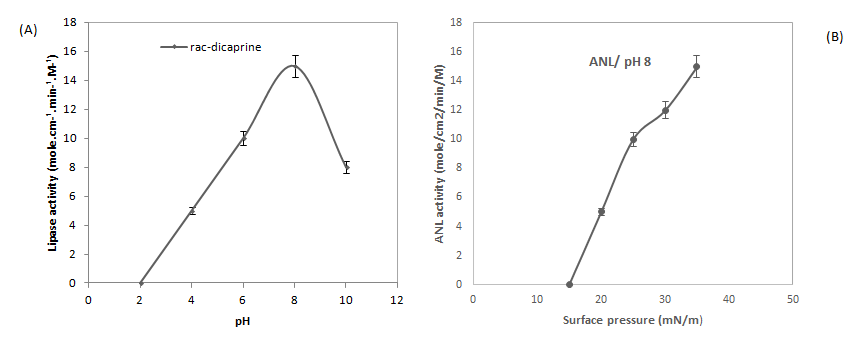
Figure 3: (A) pH dependency of the catalytic activity of ANL. Variations with pH in ANL activity using 1,2-rac-dicaprin monomolecular films. The 1,2-rac-dicaprin monolayer was spread and maintained at a constant surface pressure of 35mN/m. Assays were carried out at room temperature in a “zero order” trough (volume, 130 ml; surface, 108.58 cm2). The final enzyme concentration was 0.3 nM. The aqueous subphase was composed of various buffers that contained 150 mM NaCl, 21 mM CaCl2, and 1 mM EDTA: 10 mM glycine (pH 3-4), 10 mM sodium acetate (pH 5-6), 10 mM phosphate (pH 7), 10 mM Tris-HCl (pH 8-9), or 10 mM borate (pH 9-10). Activity is expressed as the number of moles of substrate hydrolysed by unit time and unit surface of the reaction compartment of the “zero order” trough for an arbitrary lipase concentration of 1 M. (B) Variation with surface pressure in ANL activity using pure dicaprin (diC10) isomers. Assays were carried out in a “zero-order” trough (volume, 130 ml; surface, 108.58 cm2). The final enzyme concentration was 0.4 nM ANL. Buffer: 10 mM Tris-HCl, pH 8, 150 mM NaCl, 21 mM CaCl2, and 1 mM EDTA. Activity are expressed as the number of moles of substrate hydrolysed by unit time and unit surface of the reaction compartment of the “zero order” trough for an arbitrary lipase concentration of 1 M.
ANL Kinetic Studies
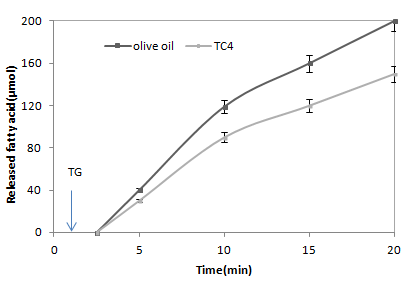
It has been established that somemicrobial lipases [16] may lost enzymatic activity in the absence of amphipathic reagent when TC4 is used as substrate. The irreversible denaturation caused by the high energy made at the tributyrin/water interface [17]. ANL is able to hydrolyse the TC4 or the olive oil emulsion without any addition of amphipathic reagent (NaDC, Triton X-100 . . .) (Figure 4). The kinetic of substrate hydrolysis remains linear for more than 20 min. Accordingly, ANL probably presents a three- dimensional structure which allows the hydrolyse of its substrate efficiently and without any denaturation at the lipide/water interface. The kinetic parameters of ANL were also studied by determining the rates of hydrolysis of different concentrations of pure TC4, TC8 or TC18. The Lineweaver- Burk curves were plotted. The apparent substrate affinity constants (KMapp.) and the maximum velocity values (Vmax) of the purified lipase were obtained and shown in Table 2. Our results showed that the KMapp. Value increased when TC4 used as substrate. In contrast, the kcat value decrease on TC8 and TC18. The deduced value of the catalytic efficiency (kcat/KMapp.) of ANL was 1226 when using TC4 as substrate. However, 4.1 or 14.7-fold decrease in catalytic efficiency was observed when using TC8 or TC18 as substrate, respectively. This result confirms that ANL hydrolyses efficiently short triacylglycerols than long-chain [18].
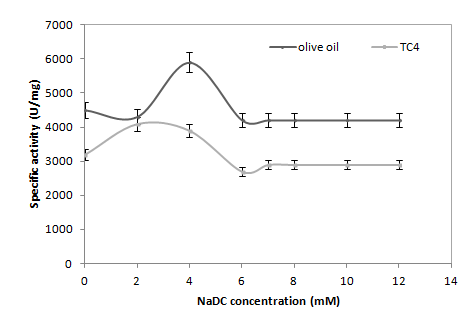
Effect of Detergents
NaDC has no inhibitory effect on the activity of lipolytic enzymes even at a high concentration (10mM) (Figure 5). This result confirms that the pure ANL is able to reach its substrate even in the presence of certain surface active agents such as bile salts. These results corroborate those obtained by Sayari A, et al. [16]. Similarly, Simons, et al. [19] showed that ANL was not inhibited by detergents such as NaDC, it can be deduced that ANL penetration power is higher than those of other microbial lipases that enables it to hydrolyse the olive oil or TC4 in presence of bile salts.
Effect of pH and Temperature on the Activity and Stability of ANL
ANL maximal activity was obtained at pH 8.0 and a temperature between 40 and 45°C. The optimum pH for ANL activity was similar to other Aspergillus Lipases [20, 21, 22]. ANL was stable at pH 5 and 8. This enzyme maintains about 100% of its activity after 30-min incubation at 60°C (data not shown). Just few fungal lipases reported in other studies presented as a thermophilic behavior [23].
Conclusion
ANL was isolated to electrophoretic purity from a culture medium. We showed that the pure enzyme, which is not a glycoprotein, is a monomer having a molecular mass of 99kDa. ANL was found to hydrolyse the short chains triacylglycerols more efficiently than the long ones. The enzyme was inactivated at 60°C and not stable at pH less than 4. Natural detergents (NaTDC, NaDC) were shown to not act as strong inhibitors of ANL as observed when bile salt was used, pH dependence of the catalytic activity of ANL was at pH 8, which confirm that ANL present one pH profile.
Acknowledgments
This work is a part of a project, and the manipulations were carried out by Dr Faouzi Laachari. This work was supported financially by Ministère de l’Enseignement Supérieur, de la Recherche Scientifique et de la Formation
des cadres-Morocco through a grant to Laboratoire de Biotechnologie Microbienne Fes.
References
-
Gumni T, Paolucci D, Bellville MP, Rios GM (2007) Enzymatic membrane reactor involving a hybrid membrane in supercritical carbon dioxide. J Membr Sci 297(1-2): 98-103.
-
Lozano P, Villora G, Gomez D, Gayob AB, Sánchez-Conesa JA, et al. (2004) Membrane reactor with immobilized Candida antarctica lipase B for ester synthesis in supercritical carbon dioxide. J Supercrit Fluid 29(1-2): 121-128.
-
Rodrigues AR, Paiva A, da Silva MG, Simões P, Barreiros S (2010) Continuous enzymatic production of biodiesel from virgin and waste sunflower oil in supercritical carbon dioxide. J Supercrit Fluid 56(3): 259-264.
-
Liaw ET, Liu KJ (2009) Synthesis of terpinyl acetate by lipase-catalyzed esterification in supercritical carbon dioxide. Bioresour Techno 101(10): 3320-3324.
-
Goddard R, Bosley J, Al-Duri B (2000) Esterification of oleic acid and ethanol in plug flow (packed bed) reactor under supercritical conditions: Investigation of kinetics. J Supercrit Fluid 18(2): 121-130.
-
Matsuda T, Kanamaru R, Watanabe K, Kamitanaka T, Harada T, et al. (2003) Control of enantioselectivity of lipase-catalyzed esterification in supercritical carbon dioxide by tuning the pressure and temperature. Tetrahedron Asymmetry 14(14): 2087-2091.
-
Wimmer Z, Zarevocka M (2010) A Review on the Effects of Supercritical Carbon Dioxide on Enzyme Activity. Int J Mol 11(1): 233-253.
-
Sajja HK (2002) Developments and trends in enzyme catalysis in nonconventional media. Biotechnol Adv 20(3-4): 239-267.
-
Hobbs HR, Thomas NR (2007) Biocatalysis in supercritical fluids, in fluorous solvents, and under solvent-free conditions. Chem Rev 107(6): 2786-2820.
-
Laachari F, El Bergadi F, El Abed S, et al. (2015) Biochemical Characterization of a New Thermostable Lipase from Bacillus pumilus Strain. Turk J Biochem 40(1): 8-14.
-
Sarda L, Desnuelle P (1958) Actions of pancreatic lipase on esters in emulsions. Biochim Biophys Acta 30(3): 513-521.
-
Bradford MMA (1976) Rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254.
-
Laachari F, El Bergadi F, El Abed S (2015) Purification and Characterization of a Novel Thermostable Lipase from Aspergillus flavus. Int J Res 2(2): 2348-6848.
-
Verger R, De Haas GH (1973) Enzyme reactions in a membrane model 1: A new technique to study enzyme reactions in monolayers. Chem Phys Lipids 10(2): 127- 136.
-
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685.
-
Sayari A, Agrebi N, Jaoua S, Gargouri Y (2001) Biochemical and molecular characterization of Staphylococcus Simulans lipase. Biochimie J 83: 863-871.
-
Horchani H, Mosbah H, Ben Salem N, Gargouri Y, Sayari A (2009) Biochemical and molecular characterisation of a thermoactive, alkaline and detergent-stable lipase from a newly isolated Staphylococcus aureus strain. J Mol Catal 56(4): 237-245.
-
Mosbah H, Sayari A, Verger R, Gargouri Y (2007) Gly311 residue triggers the enantioselectivity of Staphylococcus xylosus lipase: A monolayer study. J Colloid Inter Sci 310(1): 196-204.
-
Simons JW, Adams H, Cox RC, Dekker N, Götz F, et al. (1996) The lipase from Staphylococcus aureus. Expression in Escherichia coli, large-scale purification and comparison of substrate specificity to Staphylococcus hyicus lipase. Biochem Eur J 242(3): 760-769.
-
Ma J, Zhang Z, Wang B, Kong X, Wang Y, et al. (2006) Overexpression and characterization of a lipase from Bacillus subtilis. Protein Expr Purif 45(1): 22-29.
-
Manoj S, Kumar S, Neha S, Kannabiran K (2010) Lipase Production by Bacillus subtilis OCR-4 in Solid State Fermentation Using Ground Nut Oil Cakes as Substrate. Curr Res J Biol Sci 2(4): 241-245.
-
Shah KR, and Bhatt SA (2011) Purification and characterization of lipase from Bacillus subtilis Pa2. J Biochem Tech 3(3): 292-295.
-
Jesus MFCO, Branco RN, Santanna GLJ, Freire DMG, Silva JG (1999) Penicillium restrictum lipases: A comparative study and characterization of enzymes with different degrees of purity. Braz J Chem Eng 16(2): 113-118.
- Spectrophotometric Determination of Lanthanum (III) and Some Rare Earths with Xylenol Orange
- Introduction and Sources of Molluscicides
- Trimetazidine: An Antianginal Drug and Not Only!
- Nature Inspired Discovery and Development of Antibacterials: An Update
- Fungal Biodegradation of Polycyclic Aromatic Hydrocarbons (PAHs)
- Recent Approaches of Impurity Profiling in Pharmaceutical Analysis: A Concise Review