The Prospects of Human CD40L-Activated AntigenPresenting B Cells
B cells are considered as professional antigen presenting cells (APCs) and being used to induce the activation of cognate T cells. Because the number of primary APC from individuals’ blood are limited, in vitro activated and expanded B cells represent a good alternative APC source for studying T cell responses against pathogen or cancer. With the discoveries of T-B collaboration and T cell help for B cell activation, several groups have successfully established CD40L-based methods for generating antigen-presenting B cells. This review aims to overview current representative CD40L-based B cell culture methods with their concomitant characteristics, applications, and significances.
Introduction
Given the fact that antigen-presenting cells (APCs) express MHC molecules and costimulatory factors essential for T cell antigen recognition and activation [1], APCs are frequently used to study T cell functions and design T cell based immunotherapy against pathogens and cancers [2]. Activated B cells are considered as professional APCs, along with dendritic cells and macrophages, as they express high levels of MHC class II molecules and costimulatory molecules [3]. Primary APC numbers isolated from individuals are usually inadequate for comprehensive studies, such as TCR epitope discovery. Since B cells are more frequent than dendritic cells and macrophages in the circulation, several in vitro culture systems have been successfully adapted to promote activation and proliferation of B cells, which can be applied as the potential source of APCs [4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18].
CD40 signaling is essential for B cell activation and is induced upon binding to its ligand CD40L primarily expressed on activated T cells [19, 20]. The in vitro CD40L system for B cell activation is intentionally designed to mimic germinal centers where B cells are activated through interactions with T cells [4]. In addition to acting as a CD40L donor, T cells offer a set of B cell activation factors, known as T-cell help, such as cytokines, chemokines, and cell-cell contact [20]. Accumulating evidence reveals that stimulation by CD40L with other activators induces the activation and proliferation of B cells to function as APCs [4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18]. The CD40L-based methods for producing antigen-presenting B cells, therefore, can be considered as a powerful research tool in T and B cell studies, which bypasses the limitations of tetramer-based assays and avoids unwanted effects from using virus transformed lymphocytes or lymphoma cells.
In this review, we overview current CD40L-based culture methods on the subject of generating APCs from human primary, non-malignant, B cells for T cell studies. Following an introduction of a short history of T cell help for B cell activation, including interleukin 4 (IL-4), CD40L, B-cell activating factor (BAFF), and IL-21, recent advances inCD40L-based B cell culture systems are carefully summarized and evaluated for their applications and significances. In sum, this review aims to provide a helpful guide for researchers who are interested in finding and utilizing CD40L-based cultures of human B cells as tools in biomedical research.
T Cell Help for B Cell Activation
T and B cells are the two main cell types in the adaptive immune system. They interact closely and frequently to mount timely and efficient immune responses triggered by pathogens, and avoid unwanted immunologic effects induced by harmless antigens [21, 22]. The spatially intimate interface between T and B cells allows the contact of surface molecules required for signal transduction on the partner cells, and vice versa [23]. In 1968, Miller and Mitchell discovered T cell help to B cells [24, 25]. They showed that individual cell transfers of T cells or B cells into irradiated mice were not sufficient to induce antibody production upon immunization, whereas transfers of both T and B cells were permissive for antibody production [24]. They further demonstrated that B cells are the source of antibodies, whereas T cells are the providers of essential B cell activators [25]. Afterwards, accumulating studies provided further confirmation by testing many other antigens, which were capable of triggering antibody production [26]. Of note, exceptions exist that B cells could be activated by certain antigens in the absence of T cell help [27].
Interleukin-4
IL-4 is the first-identified help factor from T cells to B cells, discovered in 1982, more than a decade after the discovery of T to B help [28]. This soluble factor secreted from mouse EL4 thymoma cells can induce B cell proliferation in synergic with BCR agonist [28]. The discovery of that soluble growth factors of B cells are derived from other immunocytes further illustrates the importance of soluble cytokines in modulation of immune cell communication. Moreover, the Th1-Th2 paradigm was elegantly demonstrated by the biased production of antibodies in mice lacking IL-4 [29].
Kuei-Ying S and Chih-Wen P. The Prospects of Human CD40L-Activated Antigen-Presenting B Cells. J Microbiol Biotechnol, 2016, 1(2): 000111.
CD40L
CD40, a type I trans membrane protein, belonging to the tumor necrosis factor (TNF) receptor super family, was discovered in 1986 [30], whose expression is elevated on many cell types including professional APCs [31]. In 1989, Liu et al showed that CD40-CD40L interaction is an essential T cell help for B cell activation [32]; B cells were activated upon CD40 stimulation to be linked to cell division and differentiation. In 1991, Banchereau’s group established the pioneer CD40L-based culture method by using stromal cells expressing human Fc receptor combined with anti-CD40 monoclonal antibodies (the CD40 system), to enumerate B cell numbers in antibody production [4]. Banchereau’s group also evaluated the effects of IL-2, IL-4, and IL-10 on cell proliferation and antibody production of activated B cells. Later, CD40L was cloned in 1992 [33, 34], and CD40L- based methods were largely used to activate primary B cells [31]. As mentioned earlier, activated B cells express high levels of MHC class II and costimulatory factors such as CD80 and CD86, which in return, are capable of activating T cells. The in-depth understanding of CD40- CD40L signaling pathways underlying the interaction of B and T cells allows researchers to gain insight into the biology of both cell types.
BAFF
BAFF is a member of the TNF super family and identified by sequence homology search of the databases in 1999 [35]. BAFF receptors and ligands are important for B cell survival and maturation [36]. B cells bind BAFF through several receptors, such as TACI, BCMA, and BAFF- R, which are expressed on B cells undergoing maturation and differentiation from transitional B cell stage to terminal plasmacytic differentiation [37]. BAFF induces signal transduction through these receptors and promotes cell survival, APC function, and differentiation of B cells [37]. Dysregulation of BAFF expression is linked to the development of several autoimmune diseases reflects the significant roles of BAFF in B cell homeostasis [38].
Interleukin-21
IL-21 is a type I cytokine that binds to receptors composed of common-gamma chain and a private receptor [39]. IL-21 was cloned in 2000, and it is considered as the most potent cytokine capable of inducing B cell proliferation and differentiation [40]. IL- 21 drives B cells to express transcriptional factors, Copyright© Kuei-Ying S and Chih-Wen P
including BCL-6, AICDA, and BLIMP-1, known for their role in hypermutation and plasma cell differentiation [41]. In addition to enhancing proliferation of CD40L- stimulated B cells [13], IL-4 and IL-21 work synergistically to trigger class-switch recombination to IgG and the secretion of IgE from activated B cells [40, 41]. It was later demonstrated that follicular helper T cells (TFH cells), a specialized T cell subset in the germinal centers, are able to serve the major T cell help to B cells, e.g., CD40L, IL-21, IL-4, and BAFF [42].
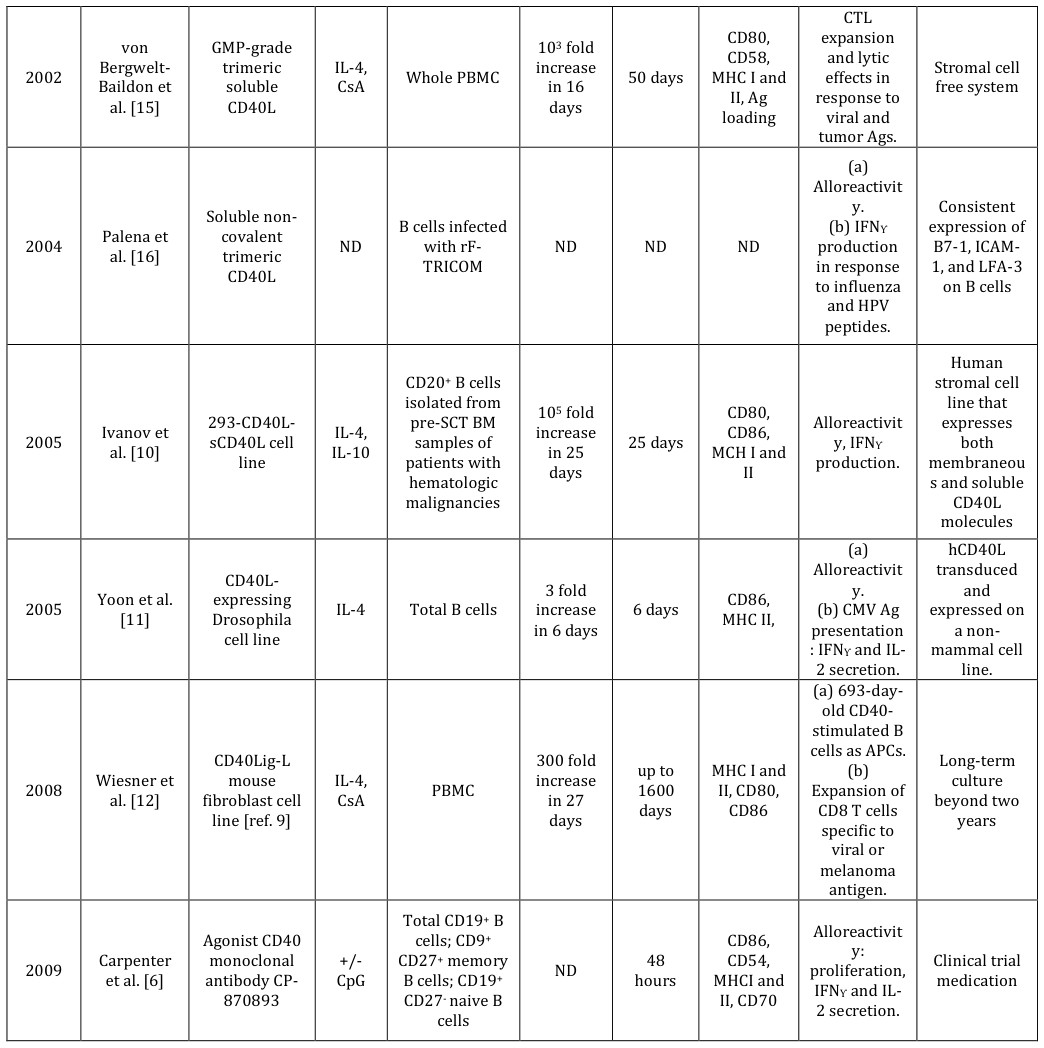
methods that have been globally employed to generate antigen-presenting B cells [4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18]. The representative works with their corresponding features are also presented. UsingCD40L-expressing cells as the source of CD40L are less expensive while stromal cells are favored because they provide much cell-cell contact than suspension cells in supporting B cell growth [43]. CD40L can be constructed in both membranous and soluble forms [10]. Although the expression levels of CD40L by stromal cells may have different effects on B cell activation and differentiation [44], CD40-CD40L signaling is an active termination signal for germinal center reaction [45]. Moreover, researchers can seek CD40L-expressing human stromal cell lines [10] or stromal free systems [4, 5, 6, 15, 16, 17, 18] if there is much concern about xenograft effects. Agonist anti-CD40 antibodies and/or recombinant CD40L molecules are available and can be used to adoptive T cell transfer for cancer immunotherapy [46].
Current CD40L-Based B Cell Cultures
As firstly proposed by Banchereau [4], CD40L-based methods were designed to mimic germinal centers where TFH cells release essential factors to activate B cells. CD40 ligation can be achieved through anti-CD40 monoclonal antibodies [4, 5, 6], soluble recombinant CD40L molecules [15, 16, 17, 18], or co-culturing with cells expressing CD40L [7, 8, 9, 10, 11, 12, 13, 14]. Table 1 presents a summary of the CD40L-based
| Efficienc | |||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| y | |||||||||||||||||||||||||||||
| Suppl | (B cell | Culture | APC | T cell | |||||||||||||||||||||||||
| Authors | CD40L | B cell | Significance | ||||||||||||||||||||||||||
| Year | ement | number | duratio | phenotyp | function | ||||||||||||||||||||||||
| (ref.) | source | populations | s | ||||||||||||||||||||||||||
| s | increase | n | e | tests | |||||||||||||||||||||||||
| in | |||||||||||||||||||||||||||||
| culture) | |||||||||||||||||||||||||||||
| 1991 | Banchereau et al. [4, 5] | Anti-CD40 mAb with hFcRII/CDw3 2 expressing mouse fibroblasts | IL-4 | Splenic or tonsillar B cells | 400 fold increase in 5 weeks | 10 weeks | ND | ND | Pioneer CD40- ligation culture | ||||||||||||||||||||
| 1995 | Schultze et al. [7] | t-CD40L cells (NIH3T3- transfectd with hCD40L) | IL-4 | Splenic B cells | ND | ND | MHC I and II, B7-1, B7-2, ICAM-1, LFA-3 | (a) Alloreactivit y: proliferation and IL-2 production. | First cell-line based CD40L system | ||||||||||||||||||||
| 1997 | Schultze et al. [8] | t-CD40L cells (NIH3T3- transfectd with hCD40L) | IL-4, CsA | Whole PBMC | 103 fold increase in 16 days; 105 fold in 61 days | 61 days | MHC I and II, CD54, CD58, CD80 and CD86 | (a) Alloreactivit y. (b) T cell proliferation, cytokine production, and cytotoxicity. (c) Halplo- mismatched T-B coculture. | Long-term culture up to 61 days. Haplo- mismatched in vitro assay |
Table 1: Description of CD40L-based cultures, cytokines, efficiency, readouts, and significances. CsA, cyclosporin A; Ag, antigen
Kuei-Ying S and Chih-Wen P. The Prospects of Human CD40L-Activated Antigen-Presenting B Cells. J Microbiol Biotechnol, 2016, 1(2): 000111.
Copyright© Kuei-Ying S and Chih-Wen P
Kuei-Ying S and Chih-Wen P. The Prospects of Human CD40L-Activated Antigen-Presenting B Cells. J Microbiol Biotechnol, 2016, 1(2): 000111.
Copyright© Kuei-Ying S and Chih-Wen P
| 2013 | Naito et al. [17] | Recombinant CD40L-Tri | IL-4 | CD19+ B cells | 30 fold increase in 14 days; 18 fold increase in day 28 | 28 days | CD80, CD86, CD83 | (a) Alloreactivit y. (b) CD8 T cells response (proliferatio n and cytokine IFNƳ production) to influenza peptide. | Another trimeric CD40L. Stromal cell free system |
|---|---|---|---|---|---|---|---|---|---|
| 2014 | Garcia- Marquez et al. [18] | Multimerized rCD40L | IL-4 | CD19+ B cells | 200 fold increase in 14 days | 14 days | CD80, CD86, HLA-DR, CD62L | Alloreactivit y | Stromal cell free system |
| 2016 | Su et al. [13] | MS40L mouse stromal cells | IL-2, IL-4, IL-21, BAFF | Naïve B cells; Memory B cells; TT-specific memory B cells | 106 fold increase in 16 days. | 16 days | MHCII, CD80, CD86 | (a) Alloreactivit y. (b) T cell proliferation against microbial Ag (HA, TT). (c) TT- specific CD4 T-cell repertoire. | The most efficient system. Expanded TT-specific memory B cells as APCs |
| 2016 | Zhang at el. [14] | 293-CD40L- sBAFF cell line | IL-2, IL-4, IL-10, CsA, CpG | B cells from PBMC | 21 fold increase in 35 days | 35 days | CD80, CD86, CD70, CD275 | (a) Alloreactivit y. (b) CTL response to HIV and melaloma antigens: proliferation, IFNƳ secretion, cytotoxicity. | Soluble BAFF secreting cell line |
Table 2: Description of CD40L-based cultures, cytokines, efficiency, readouts, and significances. CsA, cyclosporin A; Ag, antigen
Cytokines and Supplements
Among the cytokines and their related supplements, IL-4 is widely used in almost all of CD40L-based cultures since its discovery [28]. Cyclosporin A is applied in B cell culture when B cells are not directly isolated from PBMC [8, 12, 15]. Several TLR agonists, such as CpG, can be supplemented to induce B cell activation through a CD40L-independent manner [47]. IL-21 was discovered more than a decade ago and it has been recognized as the most efficient cytokine among soluble T cell help factors Kuei-Ying S and Chih-Wen P. The Prospects of Human CD40L-Activated Antigen-Presenting B Cells. J Microbiol Biotechnol, 2016, 1(2): 000111.
[39, 48]. Indeed, the addition of IL-21 was shown to promote generating large number of B cells that are capable of acting as APC [13]. The introduction of BAFF to the CD40L-based culture seems to produce beneficial effects on cell survival but not cell proliferation [13, 14].
Characteristics and Applications
Functional characteristics of B cells derived from CD40L-based cultures to be used as APCs are determined Copyright© Kuei-Ying S and Chih-Wen P
by certain representative surface markers and their ability to uptake and to present antigens to induce T cell activation. Alloreactivity assay is relatively easy to perform and is commonly used to score APC function of cultured-expanded B cells. However, since the mechanism of alloreactivity is not entirely understood [49], APC function of cultured B cells should be further confirmed by antigen presentation to autologous T cells. Meanwhile, appropriate (positive and negative) controls should always be included in testing APC function and T cell responses to facilitate accurate interpretation. The B cell sources for culture can be whole PBMC, isolated CD19+ B cells, or B cell subsets (e.g., naïve B cells, memory B cells, or selected antigen memory B cells) according to the study designs of the culture system. With the generation of enormous amounts of antigen antigen- presenting B cells, a considerable number of T cell studies have been conducted (not listed). For example, CD4 T cell differentiation and repertoire can be evaluated [13]. The application of adoptive CD8 T cells transfer is proposed as anti-HBV immunotherapy [50]. Furthermore, cognate interactions between T and B cells specific to antigens of interest can be assessed [51].
Concluding Remarks
T cells are the primary sources of membranous and soluble factors known to be essential for B activation. The CD40L-based in vitro culture system resembles in vivo germinal centers where TFH cells tightly react with B cells [41]. This review summarizes current representative CD40L-based culture methods for generating antigen- presenting B cells and these methods are widely used in many T cell studies. Moreover, the adapted culture systems can be further used to enhance APC function in order to fit the specific needs whenever necessary. Finally, with the ability to culture antigen-specific memory B cells, it is conceivably expected that cognate T and B interactions can be more easily scrutinized and to be considered as part of precision medicine.
References
-
Smith Garvin JE, Koretzky GA, Jordan MS (2009) T cell activation. Annu Rev Immunol 27(1): 591-619.
-
Sharma G, Holt RA (2014) T-cell epitope discovery technologies. Hum Immunol 75(6): 514-519.
-
Rodríguez Pinto D (2005) B cells as antigen presenting cells. Cell Immunol 238(2): 67-75. Kuei-Ying S and Chih-Wen P. The Prospects of Human CD40L-Activated Antigen-Presenting B Cells. J Microbiol Biotechnol, 2016, 1(2): 000111.
-
Banchereau J, Rousset F (1991) Growing human B lymphocytes in the CD40 system. Nature 353(6345): 678-679.
-
Banchereau J (2015) Generation of human B-cell lines dependent on CD40-ligation and interleukin-4. Front Immunol 6(55): 1-3.
-
Carpenter EL, Mick R, Rüter J, Vonderheide RH (2009) Activation of human B cells by the agonist CD40 antibody CP-870,893 and augmentation with simultaneous toll-like receptor 9 stimulation. J Transl Med 7: 93.
-
Schultze JL, Cardoso AA, Freeman GJ, Seamon MJ, Daley J, et al. (1995) Follicular lymphomas can be induced to present alloantigen efficiently: a conceptual model to improve their tumor immunogenicity. Proc Natl Acad Sci USA 92(18): 8200-8204.
-
Schultze JL, Michalak S, Seamon MJ, Dranoff G, Jung K, et al. (1997) CD40-activated human B cells: an alternative source of highly efficient antigen presenting cells to generate autologous antigen- specific T cells for adoptive immunotherapy. J Clin Invest 100(11): 2757-2765.
-
O'Nions J, Allday MJ (2004) Proliferation and differentiation in isogenic populations of peripheral B cells activated by Epstein–Barr virus or T cell-derived mitogens. J Gen Virol 85(4): 881-895.
-
Ivanov R, Aarts T, Hagenbeek A, Hol S, Ebeling S (2005) B-cell expansion in the presence of the novel 293-CD40L-sCD40L cell line allows the generation of large numbers of efficient xenoantigen-free APC. Cytotherapy 7(1): 62-73.
-
Yoon SH, Cho HI, Kim TG (2005) Activation of B cells using Schneider 2 cells expressing CD40 ligand for the enhancement of antigen presentation in vitro. Exp Mol Med 37(6): 567-574.
-
Wiesner M, Zentz C, Mayr C, Wimmer R, Hammerschmidt W, et al. (2008) Conditional immortalization of human B cells by CD40 ligation. PLoS One 3(1): e1464.
-
Su KY, Watanabe A, Yeh CH, Kelsoe G, Kuraoka M (2016) Efficient culture of human naive and memory B cells for use as APCs. J Immunol 197(10): 4163- 4176. Copyright© Kuei-Ying S and Chih-Wen P
-
Zhang Y, Gao S, Chen Y, Su L, Luo B, et al. (2016) Efficient generation of antigen-specific CTLs by the BAFF-activated human B Lymphocytes as APCs: a novel approach for immunotherapy.
-
von Bergwelt-Baildon MS, Vonderheide RH, Maecker B, Hirano N, Anderson KS, et al. (2002) Human primary and memory cytotoxic T lymphocyte responses are efficiently induced by means of CD40- activated B cells as antigen-presenting cells: potential for clinical application. Blood 99(9): 3319-3325.
-
Palena C, Zhu M, Schlom J, Tsang KY (2004) Human B cells that hyperexpress a triad of costimulatory molecules via avipox-vector infection: an alternative source of efficient antigen-presenting cells. Blood 104(1): 192-199.
-
Naito M, Hainz U, Burkhardt U, Fu B, Ahove D, et al. (2013) CD40L-Tri, a novel formulation of recombinant human CD40L that effectively activates B cells. Cancer Immunol Immunother 62(2): 347-357.
-
Garcia Marquez MA, Shimabukuro Vornhagen A, Theurich S, Kochanek M, Weber T, et al. (2014) A multimerized form of recombinant human CD40 ligand supports long-term activation and proliferation of B cells. Cytotherapy 16(11): 1537- 1544.
-
Noelle RJ, Roy M, Shepherd DM, Stamenkovic I, Ledbetter JA, et al. (1992) A 39-kDa protein on activated helper T cells binds CD40 and transduces the signal for cognate activation of B cells. Proc Natl Acad Sci USA 89(14): 6550-6554.
-
Crotty S (2015) A brief history of T cell help to B cells. Nat Rev Immunol 15(3): 185-189.
-
Tsai LM, Yu D (2014) Follicular helper T-cell memory: establishing new frontiers during antibody response. Immunol Cell Biol 92(1): 57-63.
-
De Silva NS, Klein U (2015) Dynamics of B cells in germinal centres. Nat Rev Immunol 15(3): 137-148.
-
Dustin Michael L (2014) What counts in the immunological synapse? Mol Cell 54(2): 255-262.
-
Miller JFAP, Mitchell GF (1968) Cell to cell interaction in the immune response: I. Hemolysin-forming cells in neonatally thymectomized mice reconstituted with thymus or thoracic duct lymphocytes. J Exp Med Kuei-Ying S and Chih-Wen P. The Prospects of Human CD40L-Activated Antigen-Presenting B Cells. J Microbiol Biotechnol, 2016, 1(2): 000111. 128(4): 801-820.
-
Mitchell GF, Miller JFAP (1968) Cell to cell interaction in the immune response: II. The source of hemolysin- forming cells in irradiated mice given bone marrow and thymus or thoracic duct lymphocytes. J Exp Med 128(4): 821-837.
-
Raff MC (1973) T and B lymphocytes and immune responses. Nature 242(5392): 19-23.
-
Defrance T, Taillardet M, Genestier L (2011) T cell- independent B cell memory. Curr Opin Immunol 23(3): 330-336.
-
Howard M, Farrar J, Hilfiker M, Johnson B, Takatsu K, et al. (1982) Identification of a T cell-derived B cell growth factor distinct from interleukin 2. J Exp Med 155(3): 914-923.
-
Kopf M, Le Gros G, Bachmann M, Lamers MC, Bluethmann H, et al. (1993) Disruption of the murine IL-4 gene blocks Th2 cytokine responses. Nature 362(6417): 245-248.
-
Clark EA, Ledbetter JA (1986) Activation of human B cells mediated through two distinct cell surface differentiation antigens, Bp35 and Bp50. Proc Natl Acad Sci USA 83(12): 4494-4498.
-
van Kooten C, Banchereau J (2000) CD40-CD40 ligand. J Leukoc Biol 67(1): 2-17.
-
Liu YJ, Joshua DE, Williams GT, Smith CA, Gordon J, et al. (1989) Mechanism of antigen-driven selection in germinal centres. Nature 342(6252): 929-931.
-
Armitage RJ, Fanslow WC, Strockbine L, Sato TA, Clifford KN, et al. (1992) Molecular and biological characterization of a murine ligand for CD40. Nature 357(6373): 80-82.
-
Hollenbaugh D, Grosmaire LS, Kullas CD, Chalupny NJ, Braesch Andersen S, et al. (1992) The human T cell antigen gp39, a member of the TNF gene family, is a ligand for the CD40 receptor: expression of a soluble form of gp39 with B cell co-stimulatory activity. The EMBO J 11(12): 4313-4321.
-
Schneider P, MacKay F, Steiner V, Hofmann K, Bodmer JL, et al. (1999) BAFF, a novel ligand of the tumor necrosis factor family, stimulates B cell growth. J Exp Med 189(11): 1747-1756. Copyright© Kuei-Ying S and Chih-Wen P
-
Naradikian MS, Perate AR, Cancro MP (2015) BAFF receptors and ligands create independent homeostatic niches for B cell subsets. Curr Opin Immunol 34: 126-129.
-
Mackay F, Schneider P, Rennert P, Browning J (2003) BAFF AND APRIL: a tutorial on B cell survival. Annu Rev Immunol 21: 231-264.
-
Liu Z, Davidson A (2011) BAFF and selection of autoreactive B cells. Trends Immunol 32(8): 388-394.
-
Parrish Novak J, Dillon SR, Nelson A, Hammond A, Sprecher C, et al. (2000) Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature 408(6808): 57-63.
-
Tangye SG (2015) Advances in IL-21 biology - enhancing our understanding of human disease. Curr Opin Immunol 34: 107-115.
-
Belanger S, Crotty S (2016) Dances with cytokines, featuring TFH cells, IL-21, IL-4 and B cells. Nat Immunol 17(10): 1135-1136.
-
Crotty S (2011) Follicular helper CD4 T cells (TFH). Annu Rev Immunol 29(1): 621-663.
-
Itoh K, Tezuka H, Sakoda H, Konno M, Nagata K, et al. (1989) Reproducible establishment of hemopoietic supportive stromal cell lines from murine bone marrow. Exp Hematol 17(2): 145-153.
-
Luo XM, Maarschalk E, O'Connell RM, Wang P, Yang L, et al. (2009) Engineering human hematopoietic stem/progenitor cells to produce a broadly neutralizing anti-HIV antibody after in vitro maturation to human B lymphocytes. Blood 113(7): 1422-1431. Kuei-Ying S and Chih-Wen P. The Prospects of Human CD40L-Activated Antigen-Presenting B Cells. J Microbiol Biotechnol, 2016, 1(2): 000111.
-
Bolduc A, Long E, Stapler D, Cascalho M, Tsubata T, et al. (2010) Constitutive CD40L expression on B cells prematurely terminates germinal center response and leads to augmented plasma cell production in T cell areas. J Immunol 185(1): 220-230.
-
Kornbluth RS, Stempniak M, Stone GW (2012) Design of CD40 agonists and their use in growing B cells for cancer immunotherapy. Int Rev Immunol 31(4): 279- 288.
-
Jiang W, Lederman MM, Harding CV, Sieg SF (2011) Presentation of soluble antigens to CD8+ T cells by CpG oligodeoxynucleotide-primed human naive B cells. J Immunol 186(4): 2080-2086.
-
Ettinger R, Sims GP, Fairhurst AM, Robbins R, da Silva YS, et al. (2005) IL-21 induces differentiation of human naive and memory B cells into antibody- secreting plasma cells. J Immunol 175(12): 7867- 7879.
-
Eckle SG, Rossjohn J, McCluskey J (2013) Alloreactivity. In: Zachary AA & Leffell MS (eds.), Transplantation Immunology (Methods in Molecular Biology 1034) Humana Press p: 3-39.
-
Wu C, Liu Y, Zhao Q, Chen G, Chen J, et al. (2010) Soluble CD40 ligand-activated human peripheral B cells as surrogated antigen presenting cells: A preliminary approach for anti-HBV immunotherapy. Virology Journal 7(1): 370.
-
Gonzalez NK, Wennhold K, Balkow S, Kondo E, Bölck B, et al. (2015) In vitro and in vivo imaging of initial B-T-cell interactions in the setting of B-cell based cancer immunotherapy. Oncoimmunology 4(9): e1038684. Copyright© Kuei-Ying S and Chih-Wen P
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania