Revolutionizing Livestock Breeding: The Power of Genome Editing in Cattle Farming
Big animals, including non-human primates, livestock, and dogs, play crucial roles in biomedical research and are crucial suppliers of meat and milk. Tropical regions' livestock performance has been hampered by environmental factors that promote the growth of parasites and illnesses and create heat stress in livestock. Heat stress interferes with an animal's ability to maintain homeostasis, which has an adverse effect on the meat and milk quality. In the tropical regions, a number of tactics have been used in an effort to get over these obstacles, but there are still no concrete answers in place. Biotechnologies have had a significant impact on cattle production in tropical nations during the past 20 years, including in vitro fertilization and genomic selection. The cutting-edge instrument in the cattle production toolbox is genome editing (GnEd). In breeding programs for tropical cattle, the potential to boost the genetic advantage in fewer generations through genome editing and genomic selection.
Introduction
In 2050, it is predicted that there will be 9.7 billion people on earth, with most of the expansion occurring in tropical or subtropical nations like those in Africa and Asia [1]. Food needs for this expanding population must be met without adding to the stress on land use. Pigs (20%), chickens (31%) and cattle and buffaloes (45%) are the top three sources of animal protein in the world [2]. So, greater food productivity is essential. However, a number of problems, including the environmental conditions that are essential for livestock because the majority of the countries are situated between the tropics of Cancer and Capricorn, have restricted crop and animal output in Asian and African nations. The tropics’ high temperatures and humidity have a number of negative effects on animal health, welfare, and productivity [3]. The tropical regions of Latin America also struggle with hot, humid weather, which makes it difficult to boost productivity [4]. Heat stress, which affects animal homeostasis and has an impact on the animal’s hormonal and metabolic status, is one of the major issues in the tropics. The body must change physiologically to meet this challenge, which causes a rise in heat loss and a fall in heat production [5].
Those changes come at a cost, which is typically expressed as poor milk and meat output and fertility. When high temperatures and humidity are present in vitro, lymphocytes, oocytes, and embryos exhibit direct effects of heat stress on cell viability [6]. The production of native breeds that are already suited to subtropical and tropical regions, such as the breeds of Zebu, can also be increased. These two methods can help lessen the impact of heat stress on productivity. Genetic enhancement initiatives can be used to implement both ideas. To benefit from heterosis, regional breeds can also mixed with foreign, non-adapted types [7, 8]. However, while introgression through interbreeding can transfer genes or genotypes connected to beneficial qualities in a specific trait, it can also transfer alleles of unfavorable traits that may further harm animal reproductive and efficiency.
However, cattle in the tropics face more issues than just heat stress. Many considerations must be given to ectoparasites, infections, pasture quality, and output throughout the year. Tropics are home to horn and screwworm flies, ticks, and diseases spread by ticks that cause financial losses [9]. Over the past 20 years, techniques for gene (or genome) editing have been created and enhanced. For what are known as precision breeding methods, these instruments enable the precise introduction of mutations in a specific gene. The Clustered Regularly Interspaced Short Palindromic Repeat (CRISPR) - associated nuclease 9 (CRISPR/Cas9 system) is one of the most efficient, user-friendly, and reasonably priced gene editing methods [10, 11, 12]. Cattle breeding now has more options thanks to genome editing (GnEd), which may be used to fix target alleles for monogenic traits, get rid of undesirable recessive alleles, and increase the frequency of advantageous alleles for polygenic traits [13, 14]. For instance, compared to genomic selection alone, editing of 20 loci of a desired characteristic could result in a genetic gain over 20 generations that is twice as large [15].
In fact, GnEd has the potential for alterations that in cattle would take decades to manifest. Without interbreeding, one or more alleles linked to a desired feature in a certain breed can be transferred (introduced) to another breed. Additionally, because the editing may be done in thousands of progeny from various cattle in an IVP laboratory, In one or two generations, the frequency of the desired allele or alleles can be increased in the target breed if they are already present but at a low frequency. Because of this possibility as well as its simplicity and low cost, genome editing is an effective technique for cattle in the tropics and a replacement for conventional breeding [14, 15]. For instance, genes related to a better body management of heat (heat tolerance) that are present in a tropically acclimated but low productive performance breed could be introduced to a heat-sensitive breed with high productivity performance. In a particular breed, it is feasible to add or increase the frequency of genes related to tick tolerance. In the tropics, genome editing is utilized for reasons other than animal breeding. The drought resistance and digestibility of tropical pastures can be increased to produce high-quality forages to feed animals all year round. By altering the routes from infection to the dissemination of the pathogen, resistance to infectious diseases can also be produced. It is possible to modify the rumen’s methanogen archaeas to produce less methane and so lessen the rumen’s contribution to global warming. Hence, the existing situation and future prospects for using genome editing technologies to boost cattle productivity in tropical conditions are the main emphasis of this review.
Genome Editing to Address Heat Stress Concerns
The tropics and subtropics suffer greatly from heat stress. Lower productivity causes an increase in demand for land over time; one worry is that as a result of climate change, the problem may worsen in tropical areas [16]. The Senepol, Romosinuano, Criollo Limonero, and Carora breeds of Bos taurus cattle from central and south America have been chosen for their capacity to adapt to tropical temperatures. According to a study, North America’s milk production could see losses of 1.7 billion/year in 2050 and 2.2 billion/year in 2080 as a result of global warming [17]. The effects of climate change on milk production in some Brazilian states could increase heat stress, which would get worse in both warmer and colder months. In the worst situation, severe heat could limit milk production to low productivity animals. In a place with a warm climate, it is thought that every 1 oC of temperature increase over thermal comfort may result in a loss of 1.15 kg of milk per day. Heat stress can affect the quality of milk and meat in addition to lowering performance. By reducing reproductive efficiency, which is manifested by poorer gamete and embryo viability, lower conception rates, and longer calving intervals, heat stress can also impair productivity [18, 19]. By reducing reproductive efficiency, which is manifested by lower gamete and embryo viability, lower conception rates, and longer calving intervals, heat stress can also impair productivity.
Genome Editing to Increase Quality and Milk Yield in Cattle
Milk is a staple diet for those who reside in tropical climates. For both large and smallholders, it is significant economically and socially in addition to nutritionally. While large farmers must efficiently produce milk in order to turn a profit, smallholders can milk cattle for their families’ needs while also earning some money by selling the extra to dairy enterprises. Both situations call for cows to produce milk in hot weather with a respectable efficiency. However most breeds of Bos taurus indicus struggle to produce milk effectively. When compared to Bos taurus, among other factors, the absence of genetic selection seems to be one of the main causes of low production [20]. Traditional genetic selection approaches based on progeny testing take some time to provide conclusive findings because of the large generational gaps and low intensity of selection. Nonetheless, several Bos taurus indicus breeds have genetic breeding programs that involve genomic selection. There is now a possibility to increase tropical cattle genetic improvement. Genome editing has made it possible to introduce SNPs associated with milk products into the genomes of thousands of embryos in a single generation. Because milk is a polygenic trait, increasing milk quantity or quality by genome editing will be a challenging endeavor that will depend on the discovery of SNPs using GWAS and fine-mapping studies. The introduction of many SNPs into pre-implantation embryos’ genomes could be one problem [20].
Applications of Gene Editing Tools in Farm Animals
Tandem restriction endonuclease and zinc finger- binding motifs Zinc finger nucleases are made up of FokI [21]. Fokl provides the cutting action whereas zinc finger proteins (ZFPs) are in charge of detecting and binding particular DNA sequences. The first zinc finger protein was discovered in 1983 in transcription factor IIIA, a transcription factor in Xenopus laevis oocytes [22]. A zinc finger unit is made up of about 30 conserved amino acids, and one of the most well- known DNA-binding motif types in eukaryotes is the Cys2- His2 zinc finger domain. A given DNA sequence is typically identified by more than three zinc finger domains since one ZFP unit can only recognize three DNA nucleotides. DNA- binding proteins are produced in a manner that is influenced by the modular structure of zinc finger proteins [23].
Zinc finger proteins have a modular structure that affects how DNA-binding proteins are made. In their DNA binding domain, ZFPs feature a unique surface structure that complements the DNA double helix [24]. This unique spatial structure, which resembles a finger, helps it reach into the deep groove of the DNA double helix and make contact with certain DNA bases. The positive charge of the zinc finger protein must be close to the phosphoric acid skeleton, the helix of the protein must be in the deep groove, and the joint structure between the zinc fingers must generally be stable [25]. All of these conditions must be satisfied for the zinc finger protein to bind to DNA. Afterwards, the FokI breaks the nucleotide chain following the binding site. When the target regions of two ZFNs are 6–8 bp apart, FokI of ZFNs can induce double-stranded breaks (DSBs), which then cause DNA repair processes such as error-prone non homologous end junctions (NHEJs) and rather accurate homologous directed repair (HDR). Throughout the past few decades, ZFN technology has been used for genetic testing, animal augmentation, and the development of animal models. The first ZFNs having in vitro activity were developed by Kim, et al. [26]. In 2010, a Japanese team used ZFNs to eliminate the foreign EGFP gene in pig somatic cells, successfully demonstrating the feasibility of the ZFN-KO approach in domestic animals [27]. In contrast to traditional transgenic technologies, ZFNs have shown significant advancement and potential. ZFN has good specificity and multiplies the effectiveness of genetic modification by thousands of times. Yet, as the first generation of gene editing technology, it still has some inescapable limitations. First of all, its gene editing effectiveness is still subpar. Seldom do ZFNs in porcine cells target more than 5% of the genes [28].
Second, the efficient commercial ZFN reagents are still rather expensive, and many researchers find it challenging to design and assemble the ZFN. Furthermore, the targeting activity of ZFN technology is still unknown due to the interdependence of nearby ZF motifs’ specificity. The spread and development of this technology are hampered by these shortcomings. Current research on zinc finger proteins primarily focuses on further identifying zinc finger proteins involved in the regulation and expression of genes in order to create models of how zinc finger proteins interact with DNA [29].
Transcription Activator like Effector Nucleases
TALENs have a similar structure to ZFNs [30]. They also include cleavage domains and DNA-binding domains. In addition, two TALENs are needed to cause DSBs. The DNA-binding domain of TALENs is known as transcription activator-like effector (TALE), and it was first discovered in effector proteins secreted by Xanthomonas. The DNA binding domain has 30 tandem repetitions of 33–35 amino acids, and each repeat domain specifies particular base pairs [31]. The target of TALE potentially may be any DNA sequence. Researchers linked an artificial TALE with the single-strand DNA-cleaving domain of FokI to produce the formidable weapon known as TALEN, which combines the characteristics of TALE and FokI. TALEN technology is less costly and easier to use than ZFNs. The repeat variable residues (RVDs) NN, NI, NG, and HD are used to distinguish between G, A, T, and C. Like ZFNs, TALENs can modify specific DNA sites by producing DSB-mediated NHEJ and HDR.
Nevertheless, TALEN exhibits higher gene editing efficiency and precision due to its comparatively simple structure and more accurate recognition of DNA sequence and decreased off-target danger, expense, and toxicity. Several studies have demonstrated that TALEN pairings could effectively cause knockout (KO) of target genes in a variety of cattle, with a knockout effectiveness of 20–60% [32].
ZFN has been successfully used to alter the genomes of animals; however it has limitations due to its intricate design, high cost, and lack of suitable targets. Based on the benefits of TALEN technology, numerous research organizations swiftly used it to cattle genome editing and genetic improvement [33]. Carlson, et al. [33] effectively modified the genomes of animals using TALEN, demonstrating the versatile nature of the technology. According to the study, 64% of TALENs in primary porcine cells were highly active. Direct TALEN mRNA injection into livestock zygotes caused target gene deletion in 75% of the embryos (in 29% of swine and 43- 75% of bovine). TALENs are used to successfully create MSTN-KO swine, cattle, and lamb [34].
The effectiveness of the TALEN was at least 10% in the animals, and there were noticeable alterations in the phenotypic of the muscles. Furthermore, the LDLR gene has undergone biallelic alterations using TALEN to create pig models of cardiovascular disorders. Cui C, et al. [35] studied human lactoferrin gene knock in utilizing TALENs after exploring knockout -Lactoglobulin gene in goats. This study demonstrated the potential of genetically modified sheep for use as effective mammary gland bioreactors and gene editing by TALEN-mediated HDR [35].
Clustered Regularly Interspaced Short Palindromic Repeats/CRISPR-Associated Protein
CRISPR-Cas9 may be a more effective method for improving the genetics of animals than the gene editing technologies mentioned above. CRISPRs, an acquired immune system defense against invasive DNA contaminants in bacteria and archaea, are regularly spaced short repeats. They work by DNA or RNA interference [36]. An intentionally modified type II CRISPR system is the CRISPR/Cas9 gene editing technology that is most frequently utilised. Trans- activating crRNA (tracrRNA), nuclease Cas9, and crRNA make up this system. The crRNA/tracrRNA combination directs nuclease Cas9 to cause a DSB at the target site of the crRNA-paired DNA sequence. Every 20-nucleotide sequence that is 50 bases apart from an NGG protospacer adjacent motif (PAM) sequence can be targeted by the CRISPR/Cas9 system. TALENs and CRISPR/Cas systems are simpler, more affordable, and more effective than ZFNs. Nonetheless, attention is drawn to the possible dangers of off-target and genetic variation [37]. Almost every biology lab may use the CRISPR/Cas9 system to conduct research because it is so straightforward to construct and operate. Researchers can edit particular genes using the CRISPR/Cas9 system by designing and synthesizing a short nucleic acid base on the sequence near PAM of the target gene and attaching it to the appropriate vector (such as PX330, PX459) via enzyme digestion. This method has received much research and application to enhance the nutrition, reproductive, and heredity of animals [38] (Figure 1).
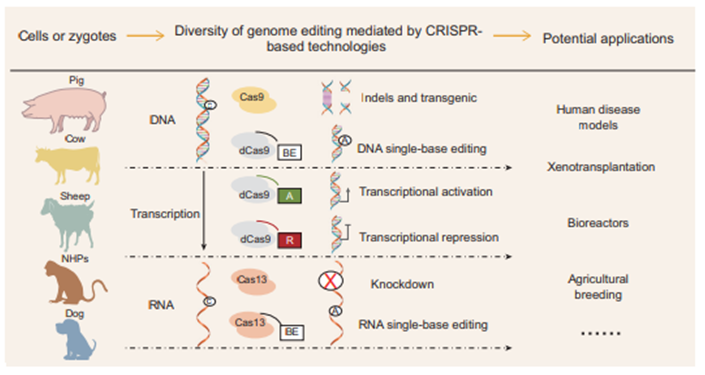
Base Editing System
Single nucleotide variations (SNVs) are the primary genetic source of 2/3 of human diseases and a significant genetic basis for phenotypic variation in animals. Consequently, it is crucial and necessary to provide an accurate and effective tool for single-base substitutions. In this situation, scientists developed the base editing system, a new CRISPR/Cas system-based target gene editing tool. Contrary to CRISPR/Cas, base-editing technologies employ nucleotide deaminases and Cas proteins that have been purposefully modified to replace bases at the target spot by producing a single incision in the double-strand. Yet, when DSBs are present, gene repairs conducted using traditional techniques (such ZFNs, TALENs, and CRISPR/Cas) typically result in a significant number of random insertions and deletions at the target locus. Point mutations are the primary cause of the majority of genetic disorders [39]. According to various base modification enzymes, the base editing systems are made up of cytosine base editors (CBE) and adenine base editors (ABE), which can achieve C-G to T-A and A-T to G-C replacements, respectively [40].
Application of Gene Editing Technology in Disease- Resistance Breeding of Livestock
Crossbreeding and selective breeding have historically been the primary ways of cattle breeding. Today’s cattle are almost entirely the result of crossbreeding and careful selection over a lengthy period of time [41]. Unquestionably, compared to the original breeds, these varieties developed through traditional breeding methods have much better production performance. On the other hand, traditional breeding methods have demonstrated to be unable of rapidly introducing or improving high-quality genes without introducing poor genes (i.e., the genes that would cause trait deficits in livestock production such infertility, susceptibility, low growth, etc.) [42]. Additionally, complex features that are challenging to see and quantify, such disease resistance traits, are difficult to choose using traditional breeding techniques. Measurement after the challenge experiment is also expensive and detrimental to the welfare and output of animals. Large-scale livestock breeding is pursuing strong disease resistance, which is antagonistic to several productive qualities.
In addition, the breeding improvement cycle is very lengthy, and several genes regulate the resistance features of many diseases. As a result, the evolution of animal breeds that are resistant has been sluggish [43]. Moreover, breeding with genome editing can quickly create novel livestock types with disease-resistance traits by directly eliminating disease susceptibility genes and pathogen receptor genes or introducing disease-resistance genes. Also, we can provide cattle with genetic features for disease resistance that are not found in naturally occurring genetic resources, leading to the development of novel livestock kinds that are not possible through conventional breeding. Swine are one of the most important livestock resources and are recognized as the greatest animal model for biomedical research and xenotransplantation due to their close physiological and genetic similarities to humans.
The FMDV virus is another economically damaging viral disease that plagues the world’s swine industry. The development of transgenic pigs that constitutively express FMDV-specific short interfering RNAs obtained from tiny hairpin RNAs is another illustration of genetic engineering for disease resistance (shRNAs). Compared to wild-type pigs, transgenic pigs exposed to the virus had no clinical symptoms of viral infection [44].
Conclusion
A genetic technology called genome editing has the potential to transform the cattle industry. There are opportunities to use this technology to improve animal performance while coping with the demands of tropical conditions. Genome editing can be carried out on the animal with the goal of introducing or increasing the frequency of one or more advantageous alleles in a specific breed or it can be carried out on the species nearby, such as tropical pastures, parasites, and microorganisms, with the goal of properly nourishing the animal and enhancing its health and quality of life. The breeding of cattle for disease resistance has benefited greatly from advances in gene editing technology, and this breeding technology will help the livestock industry experience sustainable growth. The safety and superiority of gene editing technology will be further assured with its advancement. With certainty, gene editing technology will improve animal breeding and produce better, healthier goods for all of humanity.
References
-
United Nations, Department of Economic and Social Afairs (2019) World Population Prospects 2019. Newyork.
-
Mottet A, Tempio G (2017) Global poultry production: current state and future outlook and challenges. World’s Poult Sci J 73(2): 245-256.
-
Oke OE, Uyanga VA, Iyasere OS, Oke FO, Majekodunmi BC, et al. (2021) Environmental stress and livestock productivity in hot-humid tropics: alleviation and future perspectives. J Therm Biol 100: 103077.
-
Garcia AB, Angeli N, Machado L, Cardoso FCD, Gonzalez F (2015) Relationships between heat stress and metabolic and milk parameters in dairy cows in southern Brazil. Trop Anim Health Prod 47(5): 889-894.
-
Santos MMD, Junior JBFS, Dantas MRT, Costa LLDM (2021) An updated review on cattle thermoregulation: physiological responses, biophysical mechanisms, and heat stress alleviation pathways. Environ Sci Pollut Res Int 28(24): 30471-30485.
-
Camargo LSA, Lavin TA, Adenot P, Araujo TD, Mendes VRA, et al. (2019) Heat shock during in vitro maturation induces chromatin modifcations in the bovine embryo. Reproduction 158(4): 313-322.
-
Miranda JECD, Freitas AFD (2009) Raças e tipos de cruzamento para produção de leite. Juiz de Fora MG, Agosto.
-
Camargo LSA, Saraiva NZ, Oliveira CS, Carmickle A, Lemos DR, et al. (2023) Perspectives of gene editing for cattle farming in tropical and subtropical regions. Anim Reprod 19(4): e20220108.
-
Simeão RM, Resende MDV, Alves RS, Filho MP, Azevedo ALS, et al. (2021) Genomic selection in tropical forage grasses: current status and future applications. Front Plant Sci 12: 665195.
-
Kim H, Kim JS (2014) A guide to genome engineering with programmable nucleases. Nat Rev Genet 15(5): 321-334.
-
Zhao D, Zhu X, Zhou H, Sun N, Wang T, et al. (2021) CRISPR-based metabolic pathway engineering. Metab Eng 63: 148-159.
-
Zhu Y (2022) Advances in CRISPR/Cas9. BioMed Res Int 2022: 9978571.
-
Hickey JM, Bruce C, Whitelaw A, Gorjanc G (2016) Promotion of alleles by genome editing in livestock breeding programmes. J Anim Breed Genet 133(2): 83- 84.
-
Camargo LSDA, Pereira JF (2022) Genome-editing opportunities to enhance cattle productivity in the tropics. CABI Agriculture and Bioscience 3(1): 8.
-
Jenko J, Gorjanc G, Cleveland MA, Varshney RK, Whitelaw CBA, et al. (2015) Potential of promotion of alleles by genome editing to improve quantitative traits in livestock breeding programs. Genet Sel Evol 47(1): 55.
-
Henry BK, Eckard RJ, Beauchemin KA (2018) Review: adaptation of ruminant livestock production systems to climate changes. Animal 12(s2): s445-s456.
-
Mauger G, Bauman Y, Nennich T, Salathé E (2015) Impacts of climate change on milk production in the United States. Prof Geogr 67(1): 121-131.
-
Summer A, Lora I, Formaggioni P, Gottardo F (2018) Impact of heat stress on milk and meat production. Anim Front 9(1): 39-46.
-
Zhang M, Dunshea FR, Warner RD, DiGiacomo K, Amponsah RO, et al. (2020) Impacts of heat stress on meat quality and strategies for amelioration: a review. Int J Biometeorol 64(9): 1613-1628.
-
Panetto J, Silva MV, Verneque RS, Machado MA, Fernandes AR, et al. (2021) Programa Nacional de Melhoramento do Gir Leiteiro - Sumário Brasileiro de Touros - Resultado do Teste de Progênie – Maio. Publicações e Bibliotecas.
-
Urnov FD, Rebar EJ, Holmes MC, Zhang HS, Gregory PD (2010) Genome editing with engineered zinc finger nucleases. Nat Rev Genet 11(9): 636-646.
-
Miller J, McLachlan AD, Klug A (1985) Repetitive zinc- binding domains in the protein transcription factor IIIA from Xenopus oocytes. EMBO J 4(6): 1609-1614.
-
Beerli RR, Barbas CF 3rd (2002) Engineering polydactyl zinc-finger transcription factors. Nat Biotechnol 20(2): 135-141.
-
Gaj T, Gersbach CA, Barbas CF 3rd (2013) ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol 31(7): 397-405.
-
Petersen B, Niemann H (2015) Advances in genetic modification of farm animals using zinc-finger nucleases (ZFN). Chromosome Res 23(1): 7-15.
-
Kim YG, Cha J, Chandrasegaran S (1996) Hybrid restriction enzymes: Zinc finger fusions to Fok I cleavage domain. Proc Natl Acad Sci USA 93(3): 1156-1160.
-
Watanabe M, Umeyama K, Matsunari H, Takayanagi S, Haruyama E, et al. (2010) Knockout of exogenous EGFP gene in porcine somatic cells using zinc-finger nucleases. Biochem Biophys Res Commun 402(1): 14-18.
-
Hauschild J, Petersen B, Santiago Y, Queisser AL, Carnwath JW, et al. (2011) Efficient generation of a biallelic knockout in pigs using zinc-finger nucleases. Proc Natl Acad Sci USA 108(29): 12013-12017.
-
Ates I, Rathbone T, Stuart C, Bridges PH, Cottle RN (2020) Delivery Approaches for Therapeutic Genome Editing and Challenges. Genes (Basel) 11(10): 1113.
-
Qian L, Tang M, Yang J, Wang Q, Cai C, et al. (2015) Targeted mutations in myostatin by zinc-finger nucleases result in double-muscled phenotype in Meishan pigs. Sci Rep 5: 14435.
-
Deng D, Yan C, Pan X, Mahfouz M, Wang J, et al. (2012) Structural basis for sequence-specific recognition of DNA by TAL effectors. Science 335(6069): 720-723.
-
Yang H, Wu Z (2018) Genome Editing of Pigs for Agriculture and Biomedicine. Front Genet 9: 360.
-
Carlson DF, Fahrenkrug SC, Hackett PB (2012) Targeting DNA with Fingers and TALENs. Mol Ther Nucleic Acids 1: e3.
-
Rao S, Fujimura T, Matsunari H, Sakuma T, Nakano K, et al. (2016) Efficient modification of the myostatin gene in porcine somatic cells and generation of knockout piglets. Mol Reprod Dev 83(1): 61-70.
-
Cui C, Song Y, Liu J, Ge H, Li Q, et al. (2015) Gene targeting by TALEN-induced homologous recombination in goats directs production of beta-lactoglobulin-free, high- human lactoferrin milk. Sci Rep 5: 10482.
-
Makarova KS, Koonin EV (2015) Annotation and Classification of CRISPR-Cas Systems. Methods Mol Biol 1311: 47-75.
-
Tan WS, Carlson DF, Walton MW, Fahrenkrug SC, Hackett PB (2012) Precision editing of large animal genomes. Adv Genet 80: 37-97.
-
Perisse IV, Fan Z, Singina GN, White KL, Polejaeva IA (2021) Improvements in Gene Editing Technology Boost Its Applications in Livestock. Front Genet 11: 614688.
-
Komor AC, Kim YB, Packer MS, Zuris JA, Liu DR (2016) Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 533: 420-424.
-
Landrum MJ, Lee JM, Benson M, Brown G, Chao C, et al. (2016) ClinVar: Public archive of interpretations of clinically relevant variants. Nucleic Acids Res 44(D1): D862-D868.
-
Telugu BP, Park KE, Park CH (2017) Genome editing and genetic engineering in livestock for advancing agricultural and biomedical applications. Mamm Genome 28(7-8): 338-347.
-
Bishop TF, Eenennaam ALV (2020) Genome editing approaches to augment livestock breeding programs. J Exp Biol 223(Pt Suppl 1): jeb207159.
-
Clark J, Whitelaw B (2003) A future for transgenic livestock. Nat Rev Genet 4(10): 825-833.
-
Chen Y, Cui Y, Shen B, Niu Y, Zhao X, et al. (2015) Germline acquisition of Cas9/RNA-mediated gene modifications in monkeys. Cell Res 25(2): 262-265.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania