PGPR-Mediated Defense Priming: A Sustainable Paradigm for Combating Biotic and Abiotic Stresses in Agriculture
In the face of complex biotic and abiotic stresses, modern agriculture seeks innovative solutions to ensure sustainable crop production. Plant Growth-Promoting Rhizobacteria (PGPR) emerges as powerful allies, offering a sustainable approach to fortifying plant defense mechanisms. This review delves into harnessing PGPR-mediated defense priming to combat both biotic and abiotic stresses in agriculture. Defense priming, a sophisticated mechanism acquired through exposure to primary stimuli, empowers plants to mount quicker and more resilient defense responses against subsequent challenges. PGPR induce a pre-conditioned state of heightened alertness, enabling rapid and robust defense responses upon stress encounters. This paradigm not only enhances plant resilience to pathogens and environmental stressors but also promotes sustainable practices by reducing chemical inputs. The review critically evaluates the mechanisms underlying PGPR-mediated priming, emphasizing its potential to modulate plant physiology, metabolite production, increased antioxidants enzymes, defense related enzymes activities and enhance stress tolerance. It further explores how PGPR can improve plant responses to a spectrum of stressors. This review also highlights PGPR-mediated defense priming as a cost-effective, enduring, chemical-free, and sustainable method for managing abiotic and biotic stresses in agriculture. Implementing this strategy offers effective crop protection with minimal fitness and environmental costs, even in harsh conditions.
Introduction
Amidst the accelerating pace of global warming and the persistent impacts of climate variability, the integrity of agricultural productivity hangs in the balance. These concerns are further compounded by environmental degradation, pollution, and the prevalence of biotic stresses on food crops, all of which jeopardize global food security [1, 2]. Environmental stressors can lead to substantial reductions in crop yields, ranging from 30% to 70% [3]. Both biotic and abiotic stressors inflict severe constraints on plant growth, yield, and survival. As the global population hurtles towards an anticipated 9-10 billion by 2050, the pressure intensifies on the agro-economic sector to meet burgeoning demand [2, 4]. This has increased a reliance on synthetic agrochemicals, deployed to temper stress severity, combat pathogens and pests, and amplify yields and overall crop productivity.
Current efforts are directed towards enhancing crop productivity without reliance on pesticides and fertilizers. Traditional breeding, which harnesses natural genetic diversity, has been pivotal in improving crops and strengthening their resilience against pathogens. However, conventional breeding’s lack of precision can lead to unpredictable results and require significant time and resources. On the other hand, genome editing techniques like CRISPR-Cas9 provide precise modifications but encounter regulatory obstacles and public resistance, with concerns about unintended side effects [5]. Despite their limitations, both approaches struggle with addressing complex traits and adapting to rapidly changing environmental conditions.
Plant defense priming is an innovative approach to crop protection. Various biological, physical, and chemical stimuli, as highlighted by Yang, et al. [6], can induce the primed state of the plant immune system, while beneficial interactions with root-colonizing microorganisms, as noted by Yu, et al. [7], have been identified as potential triggers for establishing this primed state. This enables primed plants to remember prior interactions with beneficial microbes, allowing them to mount quicker and more targeted defences against invading pathogens [6, 7]. This readiness, known as priming, enhances the plant’s defense mechanisms, offering more effective protection against pathogens when they attack [8]. Unlike direct defense activation, priming conserves resources by only activating defences when needed, avoiding negative impacts on plant growth and development [9, 10, 11, 12, 13, 14]. Additionally, priming provides broad-spectrum protection in challenging environments, boosting productivity with minimal fitness costs [15].
Upon receiving initial priming cues, plants undergo various physiological, transcriptional, metabolic, and epigenetic changes [9]. Some alterations in plants can lead to somatic immune memory, while others extend to reproductive tissues, forming intergenerational or transgenerational immune memory, benefiting future generations [6, 16, 17, 18]. Epigenetic modifications, such as DNA methylation and histone modifications, are crucial for establishing memory in plants [19, 20]. These changes enable plants to “remember” previous stress events and pass this information on to subsequent generations, thus supporting long-term adaptation strategies [21]. Catoni, et al. [22] discovered that priming tomato plants with β-aminobutyric acid (BABA) significantly reduced global cytosine methylation, particularly in CHH sequence contexts. This hypomethylation predominantly occurred in differentially methylated regions (DMRs) and might influence priming indirectly by affecting regulatory genes that control stress responses, rather than directly modifying defense genes. The heritability of DNA methylation marks over several generations highlights the potential of using epigenetic mechanisms to develop crops with improved stress resilience [23].
Histone acetylation and methylation have been identified as key regulators of defense priming, occurring at specific histone residues and often alongside transcriptional reprogramming due to pathogen challenges or environmental stimuli. This process results in a sustainable reconfiguration of the nucleosome structure, as highlighted by Espinas, et al. [24]. Remarkably, in Arabidopsis, researchers examined stress memory in response to hyperosmotic stress priming. This priming resulted in histone modifications such as H3K27me3, which persisted in certain genes even after ten days of normal growth, indicating that the H3K27me3 marks were passed on through mitosis [21]. Geng, et al. [25] investigated the epigenetic diversity and memory in Thlaspi arvense, an oilseed crop, under salinity stress. They found that, compared to control plants, T. arvense plants showed increased epigenetic diversity in response to salinity stress. These changes persisted in the second and third generations, suggesting that the plants retained the stress memory and could pass it on to future generations. This process, where epigenetic marks are inherited across multiple generations without recurrent stress, is known as transgenerational epigenetic inheritance. Thus, the stress tolerance resulting from epigenetic priming could be harnessed to cultivate plants with enhanced tolerance.
This inheritance of epigenetic changes can lead to intergenerational immune priming (IGIP) or transgenerational immune priming (TGIP) as shown in Figure 1 [9, 26]. Such phenomena have been observed in various crop plants, where primed plants produce resilient offspring [13, 14, 18, 27, 28]. For instance, recent studies have shown that Trichoderma-mediated priming in barley results in progeny that perform better under drought stress compared to non-primed barley. The expression of the epigenetic regulator gene HvDME was notably higher in primed barley and its subsequent generations, indicating the inheritance of priming through these epigenetic markers [20]. Another study demonstrated that primed common bean (Phaseolus vulgaris L.) with INA (2,6-dichloro-isonicotinic acid) provides resistance against Pseudomonas syringae pv. phaseolicola, with this resistance being transgenerationally transmitted via epigenetic modifications of INA-responsive
genes like PvPR1 [28]. One more research further showed that DNA methylation changes in BABA-primed potatoes confer intergenerational resistance to Phytophthora infestans, with stress memory passed through R3a promoter methylation. Similarly, Meller, et al. [29] found that in BABA- primed plants, the H3K4me2 label acts as a heritable mark for intergenerational regulation of resistance in potato.
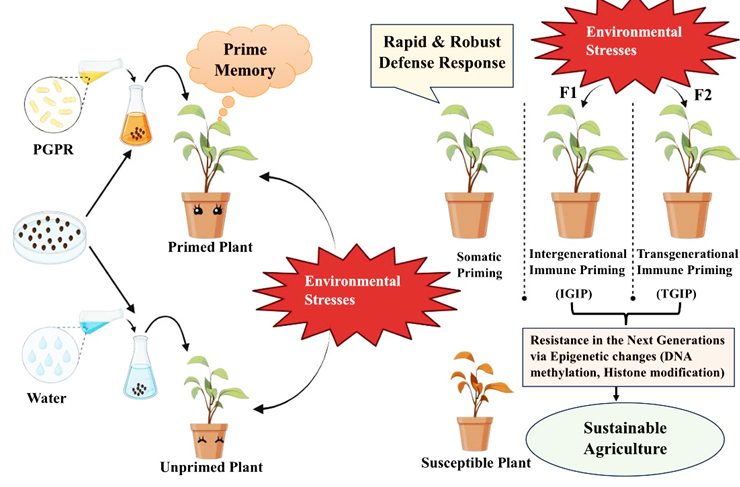
Figure 1: PGPR-mediated defense priming enhances plant resilience to environmental stresses and induces somatic memory, termed as somatic priming. Furthermore, this priming can be inherited by progeny, resulting in intergenerational and transgenerational immune priming. Epigenetic modifications, including DNA methylation and histone modification, facilitate this inheritance mechanism. Compared to unprimed plants, primed plants display heightened resilience to stresses and transmit this enhanced defense capability to future generations through epigenetic mechanisms, offering sustainable solutions in agriculture.
Recent research has demonstrated that PGPR-mediated priming in wheat against Bipolaris sorokiniana extends beyond the parent generation, establishing intergenerational immune priming. This inheritance mechanism provides sustained protection against B. sorokiniana, highlighting the potential of PGPR-based strategies for long-term disease management in agriculture [14]. Utilising beneficial microorganisms not only boosts plant defense mechanisms but also reduces reliance on chemical pesticides, promoting eco-friendly agriculture. PGPR plays a pivotal role in promoting plant growth and development through the secretion of a diverse array of modulatory compounds [30, 31].
In this context, this review aims to explore the concept of harnessing PGPR-mediated defense priming as a sustainable paradigm for combating both biotic and abiotic stresses in agriculture. We will delve into the mechanisms underlying PGPR-induced defense priming, examine its potential applications in enhancing crop resilience, and discuss its implications for sustainable agricultural practices.
Additionally, further research is needed to explore the mechanism of intergenerational and transgenerational effects of PGPR defense priming, elucidating its long-term benefits for crop protection and food security.
Efficacy of PGPR Priming for Enhancing Plant Resilience in Alleviating Environ- mental Stresses
A crucial aspect of plant defense mechanisms involves phytohormones like jasmonic acid (JA), salicylic acid (SA), and ethylene (ET), which play pivotal roles in PGPR-mediated priming to enhance plant immunity [32]. PGPR-induced defense priming and ISR rely on intricate crosstalk between signalling pathways, particularly involving JA, ethylene, and NPR proteins [33]. Understanding this network is vital for enhancing plant immunity. Perception of PGPR signals triggers cascades, activating modulating phytohormones like JA and ethylene, priming defense genes and enhancing PR protein production. This primes plants for rapid, robust responses to pathogens and provides enduring protection against diverse stresses. Additionally, PGPR-mediated ISR primes PR protein expression, linked to NPR proteins and modulation of JA and ethylene pathways [34]. JA activates defense responses against necrotrophic pathogens and herbivorous insects by promoting the expression of defense genes. SA is crucial for systemic acquired resistance (SAR), providing long-lasting protection against a broad spectrum of pathogens [35]. Ethylene works synergistically with both JA and SA to modulate defense responses, fine-tuning the balance between different hormone pathways. Certain PGPR strains can trigger ISR through the synergistic activation of JA/ ET and SA pathways [36]. For example, Bacillus thuringiensis primes tomato plants by inducing all three hormonal signaling pathways (SA, JA, and ET), thereby enhancing resistance to various pathogens [37]. Bacillus subtilis MBI600 also activates JA and SA pathways simultaneously in tomato plants to control soil-borne pathogens [38]. Similarly, Bacillus cereus AR156 can trigger immune responses against Pseudomonas syringae pv. Tomato DC3000 through ISR stimulated by both the JA/ET and SA signaling pathways [39]. In addition, B. cereus AR156 mediates priming in Arabidopsis thaliana by simultaneously activating salicylate- and jasmonate/ethylene-dependent signaling pathways [36]. NPR (Non-expressor of Pathogenesis-Related) proteins, particularly NPR1, are key regulators in the SA signaling pathway, activating the expression of PR genes and enhancing resistance. NPR proteins serve as central integrators, merging signals from SA, JA, and ethylene pathways [40]. During pathogen attack, NPR1 modulates the cross-communication between SA and JA pathways to prioritize suitable defense mechanisms [40]. Together, these phytohormones and NPR proteins form a complex network that underpins PGPR- mediated priming, enabling plants to effectively respond to and withstand environmental stresses. Furthermore, PGPR-induced defense priming not only enhances secondary metabolite production and boosts antioxidant activity but also increases osmolyte accumulation, thereby enhancing the plant’s resilience to environmental stresses (biotic and abiotic) as shown in Figure 2 [2].
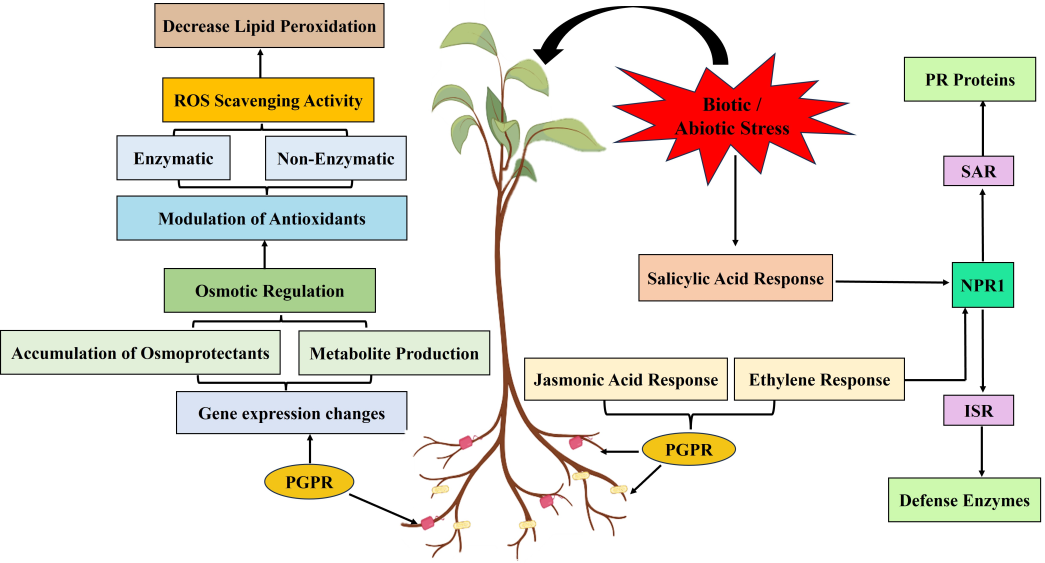
Figure 2: This schematic illustrates the signaling network underlying PGPR-induced defense priming and ISR in plants. PGPR- derived signals initiate a cascade of events, including modulation of phytohormone signaling, particularly involving JA and ethylene. This primes defense-related genes, activates defense enzymes, and increases production of PR proteins, preparing the plant for rapid defense against environmental stresses. Additionally, PGPR-mediated ISR primes PR protein expression through NPR proteins and modulates JA and ethylene pathways, facilitating streamlined defense responses. PGPR-induced defense priming also enhances secondary metabolite production and antioxidant activity, reducing oxidative stress.
Biotic Stress
Recent studies have consistently highlighted the effectiveness of plant growth-promoting rhizobacteria (PGPR) in fortifying plant defenses against diverse pathogens. For example, Sufyan, et al. [41] demonstrated that PGPR priming of chickpea seeds not only increased overall plant biomass but also decreased disease incidence caused by Fusarium oxysporum f. sp. ciceris. Likewise, Dehkian, et al. [42] showed that priming tomato plants with Bacillus thuringiensis boosted the expression of defensive genes against Fusarium oxysporum (Fol), leading to diminished disease severity. In wheat plants challenged by Puccinia striiformis f. sp. tritici (Pst), Mashabela, et al. [43] observed notable metabolic changes induced by PGPR priming, including heightened levels of phenolic compounds, indicating adaptive responses to stripe rust infection over time. Moreover, Carlson, et al. [44] employed metabolomics to reveal that PGPR-primed sorghum seedlings exhibited enhanced upregulation of defense-related metabolites against Fusarium pseudograminearum, suggesting evidence of induced systemic resistance (ISR) and a primed state in the plants. Additionally, Devi, et al. [14] underscored the protective effects of PGPR priming in wheat against Bipolaris sorokiniana, attributing this to increased levels of enzymatic and non-enzymatic antioxidants, as well as heightened activities of defense-related enzymes in primed plants compared to non-primed ones.
In a recent contribution, Tahir Ali, et al. [45] demonstrated that priming Crocus sativus with the Bacillus sp. D5 strain enhanced biomass and fortified resistance against Fusarium oxysporum infection by inducing various defense enzymes. Collectively, these studies emphasize the potential of PGPR priming as a sustainable strategy for enhancing plant defense mechanisms and reducing pathogen-induced losses in agricultural contexts.
Abiotic Stress
Due to their immobile nature, plants face a variety of abiotic stresses like drought, heat, heavy metal toxicity, and salinity, all of which hinder their growth and development [46, 47, 48]. These stresses adversely affect critical physiological processes such as photosynthesis, floral formation, seed germination, and even induce responses like stomatal closure. By 2050, it is anticipated that drought-affected crop lands will double, while water resources will diminish by thirty percent [49]. Abiotic stresses disrupt the delicate balance between reactive oxygen species (ROS) generation and scavenging, leading to the acceleration of ROS propagation and subsequent damage to crucial macromolecules and photosynthetic complexes, ultimately resulting in cell death [49]. PGPR priming enhances crop growth by activating antioxidant defense systems, regulating both enzymatic (CAT, SOD, APX, GR) and non-enzymatic antioxidants, thus shielding plants from oxidative damage [50].
The efficacy of priming techniques in enhancing drought tolerance across various crops is highlighted by recent research findings. For instance, priming maize with PGPR boosts phenolic acid flavonoids and enhances the activity of antioxidant enzymes such as SOD, CAT, and APX under drought conditions [51]. Furthermore, studies have demonstrated the effectiveness of priming in other crops. Priming Oryza sativa seeds with Bacillus pumilus SH-9 not only enhances germination metrics but also improves seedling characteristics compared to the control group, even under drought stress [52]. Similarly, priming wheat seeds with the endophytic strains B. subtilis 104 and 26D enhances growth, elevates photosynthetic pigment content, increases leaf area, and mitigates oxidative and osmotic cell damage under combined herbicide and drought stress conditions [53]. Additionally, priming wheat seeds with Bacillus subtilis 10-4 increases drought tolerance by augmenting photosynthetic pigments, relative water content, and reducing lipid peroxidation and electrolyte leakage during drought stress [54]. Moreover, priming okra seeds with Pseudomonas fluorescence enhances drought stress tolerance by maintaining relative water content, elevating the activity of both enzymatic and non-enzymatic antioxidants (e.g., AsA, GSH, SOD, CAT, APX, and GPX), and promoting metabolite accumulation, thereby improving growth under drought stress conditions [55]. In another study by Arafa SA, et al. [56], priming pea seeds with Bacillus thuringiensis MH161336 and carrot extract enhances growth, biochemical, and physiological traits. This includes increased relative water content and chlorophyll levels (a and b), up-regulation of antioxidant enzymes, higher seed yield, and reduced lipid peroxidation and reactive oxygen species during drought stress. Collectively, these findings underscore the potential of priming techniques to mitigate oxidative stress and bolster crop resilience against drought.
Soil salinity poses a significant global concern, detrimentally impacting agricultural yields in numerous countries [57]. In the realm of salinity tolerance, PGPR priming demonstrates remarkable efficacy across various crops. In peas, it modulates biochemical parameters, reducing electrolyte leakage and H2O2 levels during salinity stress [58]. Similarly, in wheat, this priming enhances water content and photosynthetic pigments while bolstering antioxidant activity under salinity stress conditions. Furthermore, PGPR priming triggers the upregulation of SOS genes (SOS1 & SOS4) in wheat during salinity stress, indicating a pathway to enhance salinity resilience [59]. Additionally, seed priming with endophytic B. subtilis strains 10–4 and 26D was found to enhance growth attributes and mitigate oxidative and osmotic damage to Phaseolus vulgaris L. plants, while also increasing lignin accumulation in plant roots under salinity stress conditions [60]. Moreover, recent research discovered that priming tomato seeds with Bacillus paralicheniformis resulted in improved germination percentage, shoot length, root length, vigor index, and dry matter production compared to the control group under salinity stress conditions [61]. Furthermore, a study revealed that seed bio-priming of maize with Bacillus spp. mitigates salt stress damage on seedlings, resulting in elongated roots. This effect is achieved by increasing the activity of catalase (CAT), peroxidase (POX), and ascorbate peroxidase (APX), while also upregulating the expression of miR160d [62]. Ultimately, these findings underscore the potential of microbial priming techniques in bolstering crop resilience to salinity stress. Another study demonstrated that priming wheat with PGPR resulted in several benefits. Primed wheat exhibited enhanced chlorophyll content and increased expression of various redox enzymes, along with the accumulation of osmolytes. This resulted in reduced membrane damage during heat stress compared to non- primed wheat plants [63]. Furthermore, another study by Jayanwita Sarkar et al. [64] demonstrated that priming wheat plants with Bacillus safensis enhanced resilience against high temperature stress. This was achieved through improvements in antioxidant enzymes, dynamic expressions of HSPs, osmolyte accumulation, and reduced chloroplast damage. Cold stress poses a significant challenge to crop development and global food security, prompting diverse molecular, physiological, and biochemical adaptations in plants across varying cold environments [65]. A study revealed that Burkholderia phytofirmans PsJN enhances Grapevine tolerance to low temperatures by upregulating stress-related genes. This priming significantly boosts the plant’s resilience compared to non-primed ones [66].
Heavy metal toxicity poses a significant threat to crop production, affecting stomata structure and function, RuBisCO activity, and disrupting photosynthesis [67, 68]. Recent research by Ahmet Direk, et al. [69] discovered that wheat seeds primed with Bacillus cereus showed enhanced growth, reduced oxidative stress markers (H2O2 and TBARS), and increased activity of antioxidant enzymes (SOD, GST, and APX). This priming led to improved yields under cadmium and arsenic stress. Similarly, a study by Aditya Banerjee & Aryadeep Roychoudhury, et al. [70] explored the priming of Oryza sativa seeds with Acinetobacter indicus, resulting in enhanced growth physiology and antioxidant capacity by activating key enzymatic antioxidants under arsenic-fluoride co-toxicity.
Conclusions and Future Perspectives
In conclusion, PGPR-mediated defense priming offers a holistic approach to mitigate both biotic and abiotic stresses in sustainable agriculture, enhancing crop resilience and reducing reliance on harmful chemicals. Future research should focus on identifying novel PGPR strains with superior priming capabilities, elucidating PGPR-plant interactions, and optimizing field application methods. Additionally, understanding the long-term impact of PGPR on plant health and soil sustainability, and integrating PGPR with other sustainable agricultural practices, will be critical for maximizing the benefits of this technology in diverse agricultural settings. By leveraging PGPR’s defense- enhancing capabilities, researchers aim to bolster plant immunity, paving the way for more enduring and resilient agriculture. Exploring transgenerational effects, elucidating molecular mechanisms, and identifying synergies with other management practices are key research avenues. Ultimately, PGPR-mediated priming presents a promising, eco-friendly tool for advancing agriculture sustainably amidst growing population demands and climate change challenges.
Author Contributions
PS and BD conceived the idea. BD, NY and PS prepared the manuscript. BD, NY and PS edited and finalised the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was funded by the Science and Engineering Research Board (SERB) under the Core Research Grant (CRG) scheme, grant number CRG/2023/001357 to PS.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
-
Goswami D, Thakker JN, Dhandhukia PC, Moral MT (2016) Portraying mechanics of plant growth promoting rhizobacteria (PGPR): A review. Cogent Food & Agriculture 2(1).
-
Mashabela MD, Piater LA, Dubery IA, Tugizimana F, Mhlongo MI (2022) Rhizosphere Tripartite Interactions and PGPR-Mediated Metabolic Reprogramming towards ISR and Plant Priming: A Metabolomics Review. Biology (Basel) 11(3): 346.
-
Etesami H, Maheshwari DK (2018) Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: Action mechanisms and future prospects. Ecotoxicol Environmen Saf 156: 225-246.
-
Fróna D, Szenderák J, Harangi-Rákos M (2019) The challenge of feeding the world. Sustainability 11(20): 5816.
-
Chawla R, Poonia A, Samantara K, Mohapatra SR, Naik SB, et al. (2023) Green revolution to genome revolution: driving better resilient crops against environmental instability. Frontiers in Genetics 14.
-
Yang Z, Zhi P, Chang C (2022) Priming seeds for the future: Plant immune memory and application in crop protection. Front Plant Sci 13: 961840.
-
Yu Y, Gui Y, Li Z, Jiang C, Guo J, et al. (2022) Induced Systemic Resistance for Improving Plant Immunity by Beneficial Microbes. Plants (Basel) 11(3): 386.
-
Luna E, Flandin A, Cassan C, Prigent S, Chevanne C, et al. (2020) Metabolomics to exploit the primed immune system of tomato fruit. HAL 10(3).
-
Mauch-Mani B, Baccelli I, Luna E, Flors V (2017) Defense Priming: An Adaptive Part of Induced Resistance. Annu Rev Plant Biol 68: 485-512.
-
Kerchev P, Meer TVD, Sujeeth N, Verlee A, Stevens CV, et al. (2020) Molecular priming as an approach to induce tolerance against abiotic and oxidative stresses in crop plants. Biotechnol Adv 40: 107503.
-
Kesel JD, Conrath U, Flors V, Luna E, Mageroy MH, et al. (2021) The Induced Resistance Lexicon: Do’s and Don’ts. Trends Plant Sci 26(7): 685-691.
-
Singh P, Tiwari M (2021) Plant Defense Priming: A New Tool for Sustainable Global Food Security: Agriculture and Innovations and Sustainability, pp: 133-153.
-
Tiwari M, Singh R, Jha R, Singh P (2022) Heritable priming by Trichoderma: A sustainable approach for wheat protection against Bipolaris sorokiniana. Front Plant Sci 13: 1050765.
-
Devi B, Tiwari M, Yadav N, Singh P (2023) Intergenerational immune priming: Harnessing plant growth promoting rhizobacteria (PGPR) for augmented wheat protection against spot blotch. Physiol Mol Plant Pathol 128: 102164.
-
Cai J, Aharoni A (2022) Amino acids and their derivatives mediating defense priming and growth tradeoff. Curr Opin Plant Biol 69: 102288.
-
Lämke J, Bäurle I (2017) Epigenetic and chromatin- based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol 18(1): 124.
-
Ramírez-Carrasco G, Martínez-Aguilar K, Alvarez- Venegas R (2017) Transgenerational defense priming for crop protection against plant pathogens: A hypothesis. Front Plant Sci 8: 696.
-
Tiwari M, Singh P (2024) Guardians of wheat: Unleashing transgenerational immune priming with Trichoderma against spot blotch. South African Journal of Botany 169: 242-254.
-
Cooper A, Ton J (2022) Immune priming in plants: from the onset to transgenerational maintenance. Essays Biochem 66(5): 635-646.
-
Tiwari M, Devi B, Sinha S, Yadav N, Singh P (2024) Intergenerational priming by Trichoderma alleviates drought stress in barley. Environmental and Experimental Botany 223: 105772.
-
Gaude AA, Siqueira RH, Botelho SB, Jalmi SK (2024) Epigenetic arsenal for stress mitigation in plants. Biochim Biophys Acta Gen Subj 1868(7): 130620.
-
Catoni M, Alvarez-Venegas R, Worrall D, Holroyd G, Barraza A, et al. (2022) Long-Lasting Defence Priming by β-Aminobutyric Acid in Tomato Is Marked by Genome- Wide Changes in DNA Methylation. Front Plant Sci 13: 836326.
-
Williams BP, Gehring M (2017) Stable transgenerational epigenetic inheritance requires a DNA methylation- sensing circuit. Nat Commun 8(1): 2124.
-
Espinas NA, Saze H, Saijo Y (2016) Epigenetic control of defense signaling and priming in plants. Front Plant Sci 7: 1201.
-
Geng Y, Chang N, Zhao Y, Qin X, Lu S, et al. (2020) Increased epigenetic diversity and transient epigenetic memory in response to salinity stress in Thlaspi arvense. Ecol Evol 10(20): 11622-11630.
-
Kotkar H, Giri A (2020) Plant epigenetics and the ‘intelligent’ priming system to combat biotic stress. Epigenetics of the Immune System, pp: 25-38.
-
Walters DR, Paterson L (2012) Parents lend a helping hand to their offspring in plant defence. Biol Lett 8(5): 871-873.
-
Martínez-Aguilar K, Hernández-Chávez JL, Alvarez- Venegas R (2021) Priming of seeds with INA and its transgenerational effect in common bean (Phaseolus vulgaris L.) plants. Plant Sci 305: 110834.
-
Meller B, Kuźnicki D, Arasimowicz-Jelonek M, Deckert J, Floryszak-Wieczorek J (2018) Baba-primed histone modifications in potato for intergenerational resistance to phytophthora infestans. Front Plant Sci 9: 1228.
-
Sharma N, Sharma G, Kour S, Chadha BS, Ohri P (2024) Unravelling the role of plant growth promoting rhizobacteria in boosting plant growth and phytoremediation of heavy metals. Applied Soil Ecology 199: 105416.
-
Gupta S, Singh UB, Kumar A, Ramtekey V, Jayaswal D, et al. (2022) Role of Rhizosphere Microorganisms in Endorsing Overall Plant Growth and Development. Re- visiting the Rhizosphere Eco-system for Agricultural Sustainability, pp: 323-353.
-
Khan N, Bano A, Ali S, Babar MA (2020) Crosstalk amongst phytohormones from planta and PGPR under biotic and abiotic stresses. Plant Growth Regulation 90(2): 189-203.
-
Salwan R, Sharma M, Sharma A, Sharma V (2023) Insights into plant beneficial microorganism-triggered induced systemic resistance. Plant Stress 7: 100140.
-
Seth K, Vyas P, Deora S, Kumar AG, Meena M, et al. (2023) Understanding plant-plant growth-promoting rhizobacteria (PGPR) interactions for inducing plant defense. In: Swapnil P, et al. (Eds.), Plant-Microbe Interaction - Recent Advances in Molecular and Biochemical Approaches. Elsevier, 2: 201-226.
-
Yadav P, Nehra A, Kalwan G, Bhardwaj D, Yasheshwar, et al. (2024) Harnessing Jasmonate, Salicylate, and Microbe Synergy for Abiotic Stress Resilience in Crop Plants. Journal of Plant Growth Regulation.
-
Afzal I, Shinwari ZK, Sikandar S, Shahzad S (2019) Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol Res 221: 36-49.
-
Dezhabad M, Taheri H, Sardrood P (2018) Bacillus thuringiensis-Mediated Priming Induces Jasmonate/ Ethylene and Salicylic Acid-Dependent Defense Pathways Genes in Tomato Plants. J Plant Mol Breed 6(2): 61-69.
-
Samaras A, Roumeliotis E, Ntasiou P, Karaoglanidis G (2021) Bacillus subtilis mbi600 promotes growth of tomato plants and induces systemic resistance contributing to the control of soilborne pathogens. Plants (Basel) 10(6): 1113.
-
Jiang CH, Huang ZY, Xie P, Gu C, Li K, et al. (2016) Transcription factors WRKY70 and WRKY11 served as regulators in rhizobacterium Bacillus cereus AR156- induced systemic resistance to Pseudomonas syringae pv. tomato DC3000 in Arabidopsis. J Exp Bot 67(1): 157- 174.
-
Li N, Han X, Feng D, Yuan D, Huang LJ (2019) Signaling crosstalk between salicylic acid and ethylene/Jasmonate in plant defense: Do we understand what they are whispering?. Int J Mol Sci 20(3): 671.
-
Sufyan M, Tahir MI, Haq MIU, Hussain S, Saeed M (2020) Effect of Seed Bio Priming With Rhizobacteria Against Root Associated Pathogenic Fungi In Chickpea. Pakistan Journal of Phytopathology 32(1): 89-96.
-
Dehkian ZK, Taheri H, Sardrood BP, Farkhari M (2024) Controlling Tomato Fusarium Wilt Disease through Bacillus thuringiensis-Mediated Defense Primining. Iran J Biotechnol 22(1): e3690.
-
Mashabela MD, Tugizimana F, Steenkamp PA, Piater LA, Dubery IA, et al. (2023) Metabolomic evaluation of PGPR defence priming in wheat (Triticum aestivum L.) cultivars infected with Puccinia striiformis f. sp. tritici (stripe rust). Front Plant Sci 14: 1103413.
-
Carlson R, Tugizimana F, Steenkamp PA, Dubery IA, Hassen AI, et al. (2020) Rhizobacteria-induced systemic resilience in Sorghum bicolor (L.) moench against Fusarium pseudograminearum crown rot under drought stress conditions. Biological Control 151: 104395.
-
Ali T, Bhagat N, Magotra S, Vakhlu J (2024) Plant Growth Promotion and Induction of Defense Response in Crocus sativus L. by Two Native Bacillus Species Against Fusarium oxysporum R1. J Plant Growth Regul 43(3): 787-806.
-
Adil S, Quraishi A (2023) A brief overview of plant abiotic stresses. NewBioWorld 5(1): 31-36.
-
Hasanuzzaman M, Nahar K, Fujita M, Oku H, Islam T (2019) Approaches for Enhancing Abiotic Stress Tolerance in Plants. CRC Press, pp: 530.
-
Ahmad M, Ali Q, Hafeez MM, Malik A (2021) Improvement For Biotic And Abiotic Stress Tolerance In Crop Plants. Biological and Clin Sci Research J 2021(1).
-
Karnwal A, Shrivastava S, Al-Tawaha ARMS, Kumar G, Kumar A, et al. (2023) PGPR-Mediated Breakthroughs in Plant Stress Tolerance for Sustainable Farming. Journal of Plant Growth Regulation, Springer.
-
Nivetha N, Asha AD, Krishna GK, Chinnusamy V, Paul S (2024) Rhizobacteria Bacillus spp. mitigate osmotic stress and improve seed germination in mustard by regulating osmolyte and plant hormone signalling. Physiol Plant 176(1): e14202.
-
Lephatsi M, Nephali L, Meyer V, Piater LA, Buthelezi N, et al. (2022) Molecular mechanisms associated with microbial biostimulant-mediated growth enhancement, priming and drought stress tolerance in maize plants. Sci Rep 12(1): 10450.
-
Shaffique S, Imran M, Adhikari A, Injamum-Ul-Hoque MD, Zainurin N, et al. (2024) Unraveling the new member Bacillus pumilus SH-9 of Bacillaceae family and its potential role in seed biopriming to mitigate drought stress in Oryza sativa. Plant Stress 11: 100318.
-
Lastochkina O, Yakupova A, Avtushenko I, Lastochkin A, Yuldashev R (2023) Effect of Seed Priming with Endophytic Bacillus subtilis on Some Physio-Biochemical Parameters of Two Wheat Varieties Exposed to Drought after Selective Herbicide Application. Plants (Basel) 12(8): 1724.
-
Lastochkina O, Garshina D, Ivanov S, Yuldashev R, Khafizova R, et al. (2020) Seed priming with endophytic bacillus subtilis modulates physiological responses of two different Triticum aestivum L. Cultivars under drought stress. Plants 9(12): 1810.
-
Pravisya P, Jayaram KM, Yusuf A (2019) Biotic priming with Pseudomonas fluorescens induce drought stress tolerance in Abelmoschus esculentus (L.) Moench (Okra). Physiol Mol Biol Plants 25(1): 101-112.
-
Arafa SA, Attia KA, Niedbała G, Piekutowska M, Alamery S, et al. (2021) Seed priming boost adaptation in pea plants under drought stress. Plants (Basel) 10(10): 2201.
-
Zilaie MN, Arani AM, Etesami H (2023) Evaluation of air pollution (dust) tolerance index of three desert species Seidlitzia rosmarinus, Haloxylon aphyllum, and Nitraria schoberi under salinity stress. Environ Monit Assess 195: 838.
-
Sapre S, Gontia-Mishra I, Tiwari S (2022) Plant Growth- Promoting Rhizobacteria Ameliorates Salinity Stress in Pea (Pisum sativum). J Plant Growth Regul 41(2): 647- 656.
-
Haroon U, Khizar M, Liaquat F, Ali M, Akbar M, et al. (2022) Halotolerant Plant Growth-Promoting Rhizobacteria Induce Salinity Tolerance in Wheat by Enhancing the Expression of SOS Genes. J Plant Growth Regul 41(6): 2435-2448.
-
Lastochkina O, Aliniaeifard S, Garshina D, Garipova S, Pusenkova L, et al. (2021) Seed priming with endophytic Bacillus subtilis strain-specifically improves growth of Phaseolus vulgaris plants under normal and salinity conditions and exerts anti-stress effect through induced lignin deposition in roots and decreased oxidative and osmotic damages. J Plant Physiol 263: 153462.
-
Parinith GR, Lakshmi S, Renganayaki PR, Nakkeeran S (2022) Alleviation of salinity stress via seed priming in tomato (Solanum lycopersicum) with Bacillus paralicheniformis. The Pharma Innovation J 11(8): 201- 206.
-
Aydinoglu F, Kahriman TY, Balci H (2023) Seed bio- priming enhanced salt stress tolerance of maize (Zea mays L.) seedlings by regulating the antioxidant system and miRNA expression. 3 Biotech 13(11): 378.
-
Sarkar J, Chakraborty B, Chakraborty U (2018) Plant Growth Promoting Rhizobacteria Protect Wheat Plants Against Temperature Stress Through Antioxidant Signalling and Reducing Chloroplast and Membrane Injury. J Plant Growth Regul 37(4): 1396-1412.
-
Sarkar J, Chakraborty U, Chakraborty B (2021) High‐ temperature resilience in Bacillus safensis primed wheat plants: A study of dynamic response associated with modulation of antioxidant machinery, differential expression of HSPs and osmolyte biosynthesis. Environ Exp Bot 182: 104315.
-
Fadiji E, Olowe OM, Akanmu AO, Ayangbenro AS, Glick BR, et al. (2024) Microbe-mediated amelioration of cold stress in plants: Mechanisms and challenges. Beneficial Microbes for Sustainable Agriculture under Stress Conditions, Elsevier, pp: 263-279.
-
Theocharis A, Bordiec S, Fernandez O, Paquis S, Dhondt- Cordelier S, et al. (2012) Burkholderia phytofirmans PsJN Primes Vitis vinifera L. and Confers a Better Tolerance to Low Nonfreezing Temperatures. Mol Plant Microbe Interact 25(2): 241-249.
-
Riyazuddin R, Nisha N, Ejaz B, Khan MIR, Kumar M, et al. (2021) A comprehensive review on the heavy metal toxicity and sequestration in plants. Biomolecules 12(1): 43.
-
Jaoudé RA, Luziatelli F, Ficca AG, Ruzzi M (2024) A plant’s perception of growth-promoting bacteria and their metabolites. Front Plant Sci 14: 1332864.
-
Direk B, Arikan-Abdulveli B, Ozfidan-Konakci C, Yildiztugay E, Uysal A (2024) Effects of Bacillus cereus on physiological and biochemical characteristics of wheat under arsenic and cadmium stress: A biological agent to reduce heavy metal stress. Plant Stress 12: 100458.
-
Banerjee A, Roychoudhury A (2023) Bio-priming with a Novel Plant Growth-Promoting Acinetobacter indicus Strain Alleviates Arsenic-Fluoride Co-toxicity in Rice by Modulating the Physiome and Micronutrient Homeostasis. Appl Biochem Biotechnol 195(11): 6441- 6464.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania