"Crocin, a Carotenoid Mitigates Acrylamide Induced Phenotypic Alterations, Oxidative Stress and Dopaminergic Neurodegeneration in Caenorhabditis Elegans: Implications for Neuropathy
Earlier we demonstrated the potential of Crocin, (CR) a well known carotenoid to attenuate experimentally induced neurotoxicity employing a Drosophila model. In the present study we sought to obtain insights on the neuroprotective propensity of CR using acrylamide (ACR) model of neurotoxicity in the nematode, Caenorhabditis elegans. We exposed age-synchronized L4 worms (both wild-type N2 and BZ555 strain to ACR (250mg/L) for 48 h in the presence or absence of CR (10μM). The protective efficacy of CR was assessed in N2 worms by analyzing phenotypic alterations (locomotory behaviour), reproductive functions (brood size and egg laying), biochemical parameters and longevity. The neuroprotective propensity was assessed by direct visualization and quantification of dopaminergic (DA) neurons in the transgenic strain (BZ555). CR treatment not only normalized reproductive functions, but significantly alleviated ACRinduced oxidative stress and mitochondrial dysfunction. Further, CR restored the activity levels of acetylcholinesterase, dopamine levels and markedly reduced the dopaminergic degeneration. CR also extended the lifespan in N2 worms and markedly enhanced the expectancy of short-lived mutant worms (MEV-1). Collectively, these experimental findings reveal that CR significantly attenuates the dopaminergic neurodegeneration and associated physiological/ biochemical dysfunctions induced by ACR in C. elegans. Based on these data we propose that Crocin may be an effective therapeutic adjuvant in the management of oxidative stress mediated neurodegenerative conditions.
Introduction
Crocus sativus L. commonly known as saffron is cultivated in many countries such as Iran, India, Italy, Spain and Greece. Commercially available saffron is the dried red stigmas of the flower. The main bioactive constituents of saffron are crocins, picrocrocin, and safranal [1]. Saffron extract or its active constituents have been demonstrated to have diverse biological properties such as antioxidant, anti-inflammatory and neuroprotective effects [2, 3, 4]. Crocins (CR) are the glucosyl esters of crocetin, which are water-soluble carotenoids. CR is responsible for its characteristic colour of saffron and numerous studies have attributed its antioxidant property to be responsible for the protective efficacy in a wide range of diseases [5]. Both i_n vitro_ and in vivo studies have demonstrated the cytoprotective, anti-inflammatory and anti-oxidant effects of CR on nerve cells [6, 7]. Several recent reports suggest the potential of CR to improve cognitive power and to reduce anger and depression [8, 9].
Acrylamide (ACR), a well-documented human neurotoxin is a commonly used chemical for various industrial applications and gel electrophoresis in laboratories [10]. ACR is a water soluble monomer, formed in carbohydrate-rich foods when they are prepared in high temperature. In recent times, ACR has gained considerable attention owing to its formation during heat processing of carbohydrate rich foods (eg., French fries, chips). Compelling evidence from different experimental models indicate its toxic effects leading to neuropathic signs. ACR monomer is known to affect both central and peripheral nervous system and impacts both sensory, motor and cognitive functions [11, 12, 13, 14]. Various undesirable effects of ACR including reproductive toxicity, carcinogenicity and neurotoxicity have been described [15, 16, 17]. ACR also induces metabolic alterations, genotoxicity and mutagenicity [18, 19]. Industrial exposure of humans to ACR results in neurological symptoms, including ataxia, skeletal muscle flaw, numbness of hands/ feet, neuropathy and Parkinsonism [20]. In vivo and in vitro studies have shown that ACR alters dopamine levels by affecting the dopaminergic system [21, 22].
Dopaminergic neuronal loss, motor impairment, formation of lewy bodies, increased oxidative stress and mitochondrial dysfunction are the characteristic features of Parkinson’s disease (PD) [23]. The advantages of employing the nematode Caenorhabditis elegans (C. elegans) to investigate the association between oxidative stress and DAergic neurodegeneration are well known [24]. The simplicity of C. elegans to high-throughput genomic, proteomic and drug screening approaches make this organism a striking option for recognizing new curative compounds for PD and understanding their mechanism of action. The simple nervous system of C. elegans contain only 302 neurons (out of 959 somatic cells) in adult hermaphrodites and various neurons are categorized as sensory, interneuron, or motor neurons. It has 8 DA neurons, with completely mapped neuronal networks [25]. C. elegans is extensively used for studying varying cellular pathways because of the availability of genetic, behavioural, biochemical and physiological endpoints associated with normal aging and neurodegeneration including PD [26]. Since DA neurons can be directly visualized through the expression of a fluorescent protein (Green fluorescent protein (GFP), this model has also been employed to induce DA neuron degeneration utilizing chemical neurotoxins to understand the mode of action of therapeutic compounds [27, 28].
Recently we demonstrated the neuroprotective effects of Saffron extract and CR in a Drosophila model of Parkinsonism [29]. In this study, utilizing C. elegans as a model system, we investigated the neuroprotective efficacy of CR focusing on the aspects related to its potential to abrogate ACR mediated neurotoxic outcome. Initially, we determined the potential effects of CR in worms and recapitulated the neurotoxic effect of ACR. Further, employing a co-exposure paradigm, the efficacy of CR to alleviate ACR-induced locomotor and physiological effects, oxidative impairments, cholinergic function and dopamine levels were determined. The neuroprotective potential of CR was assessed by quantifying the extent of neuronal degeneration in the transgenic strain (BZ555). Furthermore, we also investigated its ability to extend the longevity employing both wild and short lived mutants (MEV-1).
Materials and Methods
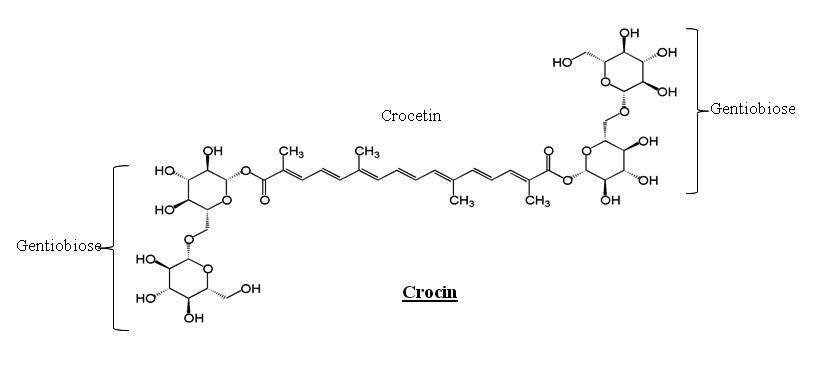
Crocin (CR, Figure 1), Acrylamide (ACR), Dopamine (DA), and other fine chemicals were procured from Sigma
Chemical Co. (St. Louis, MO, USA). All other chemicals used were of analytical grade. C. elegans, wild-type strain (N2), BZ555 strain (GFP tagged in dopamine neuronal stroma and processes), a mutant strain of C_. elegans_ (MEV-1) and Escherichia coli (OP50-uracil auxotroph) were obtained from the Caenorhabditis Genetics Center (CGC, Minneapolis, MN, USA) which is funded by the National Centre for Research Resources (NCRR).
C. elegans Maintenance and Treatment
C. elegans were maintained on Nematode Growth Medium (NGM) seeded with E. coli OP50 and sustained at 20◦C. Age-synchronized worms were obtained by the alkali-bleaching method [30].
Crocin (CR) Concentrations: Effect on Endogenous Levels of Oxidative Markers
For this study, L4 stage worms were exposed to varying concentrations of CR (5, 10 and 20µM) for 48h and the effect on locomotor function was evaluated. Further, the effect of CR on endogenous levels of biochemical markers of oxidative stress was determined.
Selection of ACR Concentration and Exposure of Worms
In a preliminary study, worms were exposed to ACR at varying concentrations (75-250mg/L) in K-medium for 48h and induction of locomotor deficits were assessed. Further we determined the impact of ACR exposure on selected biochemical markers of oxidative stress. Based on these results, a concentration of 250 mg/L was selected for further experimentation in the co-exposure paradigm.
ACR Exposure - Recapitulation of ACR Induced Effects in Worms
The basic experimental design comprised of exposing L4 stage worms to ACR for a period of 48 h. Subsequently, worms were washed thrice with K-medium and subjected to various studies. Assessment of behavioural phenotypes consisted of thrashing behaviour, pharyngeal pumping and locomotor movements. Biochemical analysis comprised of quantification of oxidative stress markers, activity levels of antioxidant enzymes, mitochondrial enzymes, cholinergic function and dopamine content.
Protective Effect of CR Treatment Against ACR- Co Exposure Paradigm
Modulation of phenotypic and biochemical alterations in N2 worms: In order to assess the modulatory effect of CR on ACR-induced phenotypic alterations, a co-exposure paradigm was employed. L4 worms were exposed to sublethal concentration of ACR (250mg/L) and CR (10µM) for 48 h in 6-well tissue culture plate. Following the exposure period, worms were washed thrice with K-medium and subjected to select physiological measurements and processed further for biochemical assays.
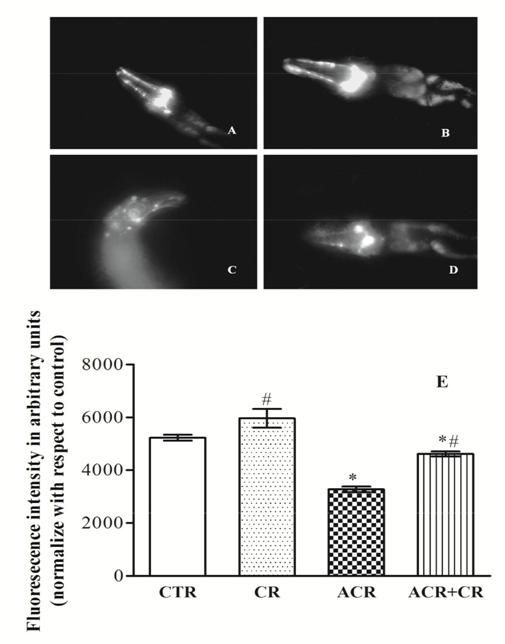
Neuroprotective effect of CR: Analysis of neuronal degeneration in BZ555: Strain Synchronized L4 stage of BZ555 (egls1 [dat-1p: GFP] i.e., bright GFP observable in Dopamine neuronal soma and processes) were exposed to ACR and CR at the concentrations stated above for 48 h. After the exposure period, the worms were washed thrice with K-medium to dispose of bacterial debris. Microscopy of living young adults was executed by introducing the worms on glass slides with a small aliquot of 100 mM sodium azide (utilized as an anesthetic) and GFP was visualized with a fluorescence microscope. All images were developed with Image J software for quantifying the green pixel number. Two independent experiments were conducted for each dose with at least 10 worms/per exposure group. Potential of CR to extend Lifespan of worms exposed to ACR: In this study, we examined the effect of CR on the lifespan of worms exposed to ACR employing both wild type (N2) and short lived mutant (MEV-1) strains employing a co-exposure paradigm. Worms (n=20 ± 1) from both control and ACR exposed groups after 48h were transferred to a 24-well plate containing K-medium with OP50 and 5-fluoro-2-deoxyuridine (FUdR: 50 µM). The worms were maintained at 20◦ C and checked daily for survivability until all the worms died [31].
Methods of Analysis
Analysis of behavioural response: Following exposure to ACR, worms were washed in K-medium and subjected to the behaviour analysis as briefly described below. Thrashing: This study was conducted by placing worms in K-medium and counting the number of tangential swimming movements (thrashes min-1), thereby measuring a significant feature of locomotion. The effects of toxin on the locomotion of worms were quantified as described previously [32]. Pharyngeal pumping: Pharyngeal pumping is defined as the backward grinding movements in the terminal bulb of the worm in the presence of Escherichia coli OP50. To assay pumping on bacteria, pumps were counted for 30 s. At least 5 worms per group were analyzed for pumping behaviour and the results pooled [33]. Locomotion: We analyzed rates of locomotion behaviour by measuring the incidence at which young adult animals initiated body bends. NGM plates with thin lawns of OP50 bacteria were used as the stage for the worms to perform locomotory behaviour. We counted the number of body bends by direct observation for a total of 30 s. A body bend was counted each time a bend reached a maximum just posterior to the pharynx, either on the dorsal or ventral side of the animal [34]. Assessment of physiological responses Lifespan: L4 worms (20 ± 1) were transferred to a 24- well plate containing K-medium with OP50 and 5-fluoro- 2-deoxyuridine (FUdR: 50 µM). The worms were maintained at 20
- C and checked daily for survivability until all the worms died [31]. Egg laying: About ten worms picked from each treatment group were shifted to NGM plates. The number of eggs laid by every worm during 1 h after exposure period was counted [35]. Brood size: A single worm was picked and relocated to 12-well tissue culture plate containing 1 ml K-medium, with OP50. The plates were incubated at 20
- C for72 h. After the incubation period, the worms were washed, pelleted and the progeny was counted under the dissecting microscope [36]. Assays were repeated three times in triplicate. Biochemical estimations Measurement of ROS generation: Reactive oxygen species (ROS) produced was evaluated using dihydro dichlorofluorescein diacetate, a non-polar compound that after conversion to a polar derivative by intracellular esterases, can swiftly react with ROS to form the highly fluorescent compound, dichlorofluorescein in [37]. In brief, an aliquot equivalent to 0.1 mg protein was incubated in Locke’s solution (pH 7.4; NaCl – 154 mM, KCl – 5.6 mM, NaHCO3– 3.6 mM, HEPES – 5 mM, CaCl2 – 2 mM and glucose – 10 mM) containing 5 μM DCFDA for 30min at room temperature and the fluorescence was considered at excitation and emission wavelengths of 480 and 530 nm, respectively. The ROS generation was projected from a DCF standard curve and expressed as pmol DCF/min/mg protein. Measurement of HP: The levels of hydroperoxides (HP) were determined based on oxidation of xylenol orange reconciled by ferrous ions [38]. An aliquot of homogenate was added to 1 ml of FOX reagent (250 μM Ferrous ammonium sulphate; 100 μM sorbitol; 25 mM H2SO4; 100 μM xylenol orange), and incubated for 30 min at room temperature. The absorbance was read at 560 nm and expressed as nmoles hydroperoxides (HP) /mg protein. Estimation of Reduced glutathione (GSH) and total thiols: The levels of reduced glutathione were established by following o-pthalaldehyde fluorescence [39]. An aliquot of the sample (0.1 mg protein) was deproteinized by adding formic acid (0.1 M) and subsequent centrifugation (10,000 rpm, 10 min). The supernatant was mixed with alkaline formaldehyde (8% in 20 mM Na2HPO4) and incubated (45 min) with sodium phosphate buffer (0.1 M, pH 8.0, containing 5 mM EDTA, 100 mg/ml o-pthalaldehyde). Reduced glutathione levels were read (excitation – 345; emission – 425) against a standard curve and expressed as μmol GSH/mg protein. The total thiols levels were determined following a previously described method with slight modification [40]. Activities of antioxidant enzymes: Superoxide dismutase (SOD) activity was calculated indirectly by observing the inhibition of quercetin auto-oxidation [41]. In brief, to a reaction mixture of phosphate buffer (0.016 M, pH 7.8, containing TEMED-4 mM and EDTA- 0.04 mM) quercetin (50 μM) was added and the rate of its auto oxidation was monitored at 406 nm for 3 min. The ability of the test sample to inhibit quercetin oxidation by 50% is defined as one unit of the enzyme and activity expressed as units/mg protein. Catalase activity was measured by adding an aliquot of cytosol (0.25 mg protein) to phosphate buffer (0.1 M, pH 7.4, containing H2O2 – 10 mM) [42]. The collapse of H2O2 was monitored at 240 nm and activity expressed as nmol substrate/ min/mg protein (€– 44.2/mM/cm). Thioredoxin reductase (TR) activity in the test sample was estimated by monitoring the reduction of DTNB at 412 nm, in a potassium phosphate buffer (0.1 M, pH 7.0, containing in mM: EDTA – 10, NADPH – 0.2) [43]. The activity was expressed as nmol substrate reduced/min/mg protein. Glutathione-S- transferase (GST) activity was quantified by scrutinizing the conjugation of glutathione to CDNB at 340 nm [44]. The reaction was started by adding a cytosolic aliquot (0.01 mg protein) to the phosphate buffer (0.1 M, pH 6.5 containing in mM: EDTA – 0.5, CDNB – 0.075, GSH – 0.05). The increase in the optical density at 340 nm was recorded for 3 min and the activity expressed as nmol conjugate formed/min/mg protein. Determination of acetylcholinesterase (AChE) activity: AChE activity was quantified according to a standard method [45]. The reaction was initiated by the addition of acetylthiocholine iodide (ATCI, 1.95 mM) to the phosphate buffer (0.1 M, pH 8.0) containing DTNB (2.5 mM) and cytosolic sample (0.01 mg protein). The change in absorbance was monitored at 412 nm for 3 min. The enzyme activity was expressed as nmol substrate hydrolysed/min/ mg protein. Analysis of dopamine concentrations by HPLC: After the exposure period, the worms were washed thrice with K-medium, pelleted and then homogenized in buffer A (50 mM Tris–HCl buffer pH 7. 4 + 0. 1% Tween 20) followed by centrifugation at 10,000 rpm for 10 min at 4
- C. The supernatant was used for dopamine quantification after filtration through 0.25m nylon filters. The quantification was done by high-performance liquid chromatography (Shimadzu HPLC) with an electrochemical detector (ECD) consisting of a high-pressure isocratic pump, a 20 µl sample injector valve, C18 reverse phase column and electro chemical detector. Retention time (RT) for dopamine peak was between 11.9and 13.3 min of the run time. Dopamine (DA) standard was used to obtain a standard curve and the DA content in the control and experimental worms were calculated and DA was expressed as pg/mg protein [46]. Determination of mitochondrial enzyme activity: NADH–cytochrome C reductase activity was quantified following the reduction of cytochrome C by the sample (0.1 mg protein) in phosphate buffer (0.1 M, pH 7.4, having NADH– 0.2 mM, KCN – 1 mM) [47]. The decrease in absorbance was monitored at 550 nm for 3 min and activity was expressed as nmol cytochrome-c reduced/min/mg protein. The MTT reduction assay was performed by incubating the mitochondrial aliquot (0.015 mg protein) in the mitochondrial re-suspension buffer (containing sodium succinate – 20 mM, MTT- 75 mg/ml) for 60 min at 37oC. The formazan formed was suspended in SDS reagent (pH 7.4, 10%-SDS, 45%-DMF), measured at 570 nm and expressed as OD/mg protein [48]. Measurement of ADP/ATP ratio: ADP/ATP ratio was quantified by ApoSENSOR™ ADP/ATP Ratio Bioluminescent Assay Kit (from Biovision Catalog #K255- 200) according to the manufacturer’s instruction. This kit utilized bioluminescent detection of the ADP and ATP levels for a rapid screening of apoptosis, necrosis, growth arrest, and cell proliferation simultaneously in cells. The assay utilized the enzyme luciferase to catalyze the formation of light from ATP and luciferin, and the light was measured using a luminometer or Beta Counter. ADP level was measured by its conversion to ATP that is subsequently detected using the same reaction in a 96 well plate. Protein estimation: Protein concentrations were determined by incubating an aliquot of the sample with Folin–Ciocalteau’s phenol in an alkaline medium and measuring the optical density at 750 nm using a UV–VIS spectrophotometer [49]. The amount of protein was enumerated using bovine serum albumin as the standard.
Statistical Analysis
Every experiment was repeated at least thrice with three replicates each. Mean and standard error (SE) were evaluated for all parameters, and the results were expressed as mean SE. The data were analyzed employing one-way ANOVA followed by the Tukey’s multiple comparison tests. The longevity data were subjected to Kaplan–Meier survival analysis to prepare survival curves and the data were compared with untreated control worms. Values ≤ 0.05 were considered as significant. All the calculations were carried out by Graph pad prism software.
Results
Effect of Crocin (CR) Treatment
Worms maintained in K-media enriched with CR (5- 20µM) for 48h exhibited no locomotor dysfunction or lethality (data not shown). However, CR treatment (48h) significantly influenced the basal levels of some important biochemical parameters. CR decreased the levels of ROS, increased GSH levels, enhanced GST activity and reduced the AChE activity levels in worms. All further experiments were conducted employing CR at concentrations of 5- 10µM.
Recapitulation of ACR Effects in Worms
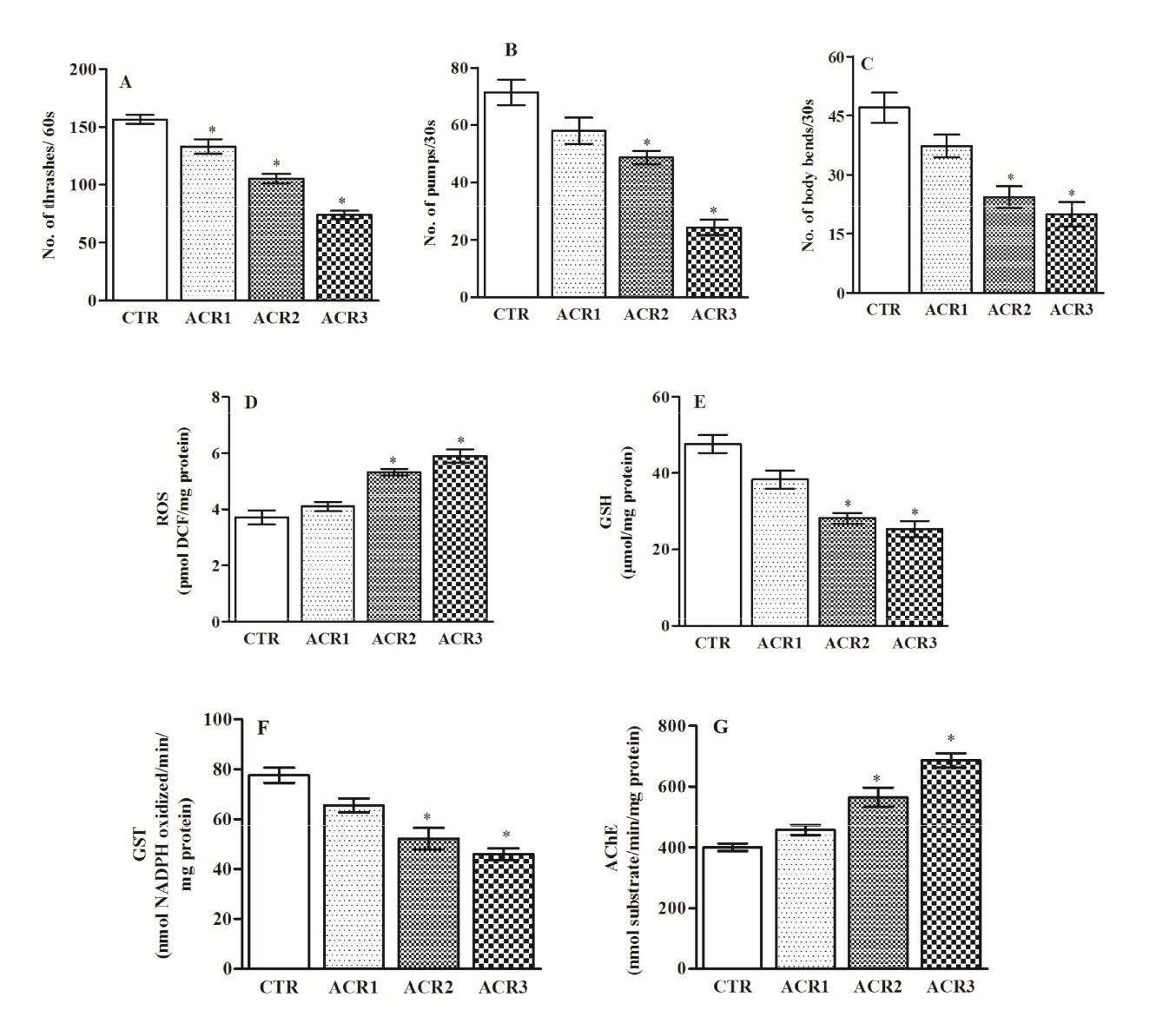
Worms exposed to different sublethal concentrations of ACR (75-250mg/L) demonstrated a concentration- dependent alteration on locomotor behaviour. While the lowest concentration of ACR had no significant effect on worms, at higher concentrations (150 and 250mg/L) it caused a significant reduction in thrashing behavior, pharyngeal pumping and locomotion Figures 2A-C. Further, ACR induced significant oxidative stress as evidenced by elevated ROS levels, decreasing GSH levels, diminished GST activity levels and enhanced AChE activity Figures 2D-G. Based on these, a concentration of 250mg/L of ACR was used for further studies to assess the protective efficacy of CR.
Modulatory Effect of CR on Behaviour Parameters of Worms Exposed To ACR
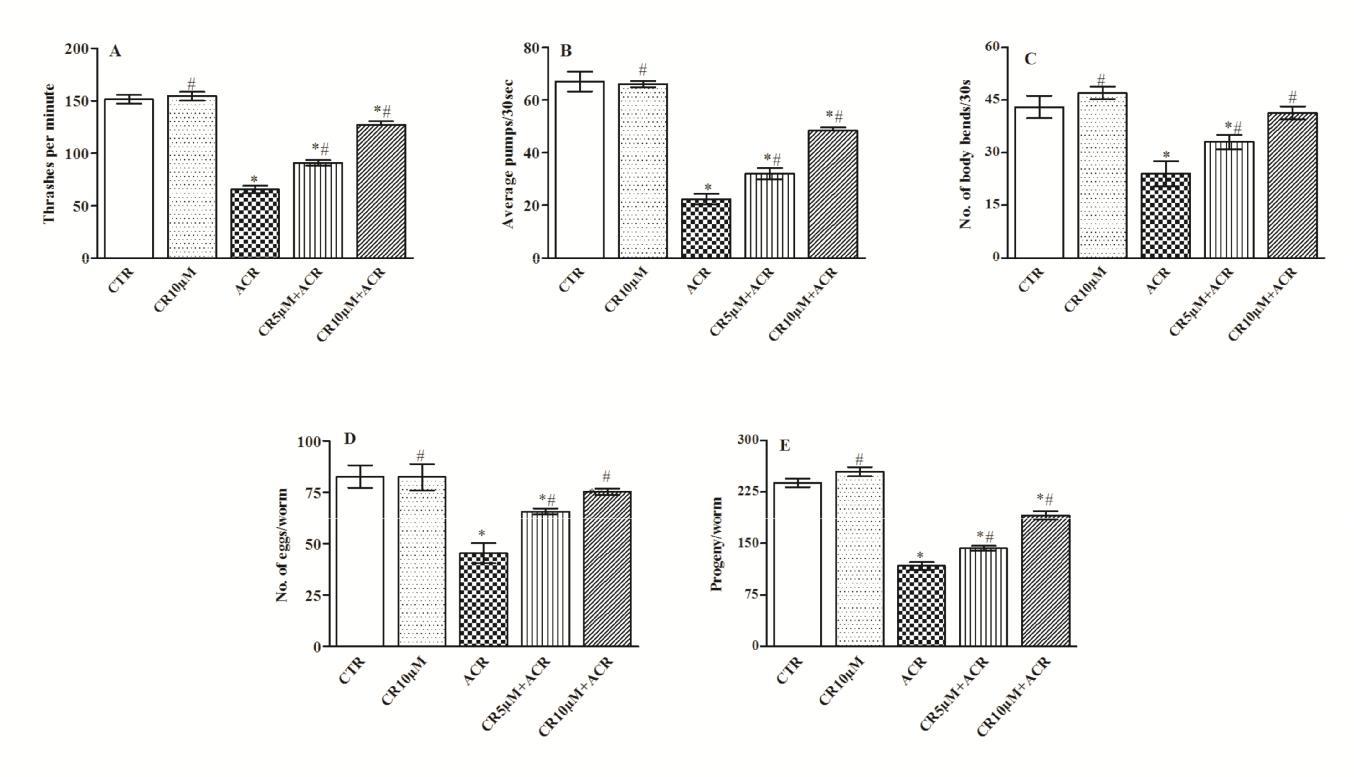
ACR exposure caused a significant decrease in the locomotor activity of worms. While ACR reduced the thrashing movements (by 57%), body bending (by 44%), and pharyngeal pumping was markedly (by 67%) diminished. Interestingly, in the co-exposure paradigm, CR treatment significantly improved all the three behaviour parameters. While the thrashing behaviour and locomotion was significantly restored (by 28-48%) Figure
3A, higher degree of restoration (30-54%) was evident with pharyngeal pumping Figures 3B & C.
Effect of CR on Egg Laying and Brood Size of Worms
Worms exposed to ACR showed a significant decrease in egg laying (45%) and brood size (51%), which were normalized by CR treatment in a concentration- dependent manner Figures 3D & E.
Protective Efficacy of CR on Redox Status and Antioxidant Enzyme Levels
The effect of ACR exposure on markers of oxidative stress and antioxidant enzyme levels were quantified in worms. ACR caused a significant increase in ROS (56%) and HP (63%) levels. While a lower concentration of CR offered only a marginal protective effect, CR at higher concentrations significantly reduced both ROS (40%) and HP (46%) levels in worms Table 1. Further, the levels of reduced GSH and total thiols which were significantly (40-
45%) depleted among worms exposed to ACR, were restored markedly with CR treatment Table 1. Furthermore, ACR exposure caused significant reduction in the activity levels of enzymatic antioxidants viz., SOD, catalase, TR and GST and CR treatment attenuated this effect as evident with enhancement in the levels of the enzymic antioxidants Table 2.
Effect of Crocin supplementation on oxidative markers, GSH and total thiols against acrylamide toxicity in C. elegans.
| Group | ROSa | HPb | GSHc | TSHd | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTR | 2.64±0.28 | 0.96±0.01 | 45.09±3.17 | 22.35±1.07 | |||||||||
| CR (10µM) | 2.21±0.4# | 1.03±0.03# | 46.33±2.04# | 29.49±0.57*# | |||||||||
| ACR (250mg/L) | 5.99±0.18* | 2.57±0.01* | 24.87±2.03* | 13.26±0.59* | |||||||||
| CR(5 µM)+ACR | 5.16±0.07*# | 2.18±0.12*# | 26.99±0.88* | 15.73±0.47*# | |||||||||
| CR(10 µM)+ACR | 3.15±0.22*# | 1.38±0.03*# | 33.73±1.13*# | 18.58±0.50*# |
Table 1: Values are mean ± SE. Data analyzed by one-way ANOVA followed by Tukey’s test (p < 0.05); *= significantly different com
Modulatory effect of Crocin supplements on the activity of antioxidant enzymes against acrylamide toxicity in C. elegans.
| Group | SODa | CATb | TRc | GSTd | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTR | 49.52±1.67 | 23.47±1.04 | 27.86±1.32 | 78.4±2.57 | ||||||||
| CR (10µM) | 66.86±2.03*# | 23.36±0.66# | 30.81±0.80# | 87.93±0.87*# | ||||||||
| ACR (250mg/L) | 29.01±0.74* | 10.36±0.65* | 16.26±0.62* | 47.43±0.91* | ||||||||
| CR(5 µM)+ACR | 31.45±0.35*# | 11.40±0.35* | 18.75±0.4*# | 52.88±0.86*# | ||||||||
| CR(10 µM)+ACR | 39.60±1.45*# | 17.23±0.55*# | 23.34±0.45*# | 69.93±2.16*# |
Table 2: Values are mean ± SE. Data analyzed by one-way ANOVA followed by Tukey’s test (p < 0.05); *= significantly different com
Effect on Mitochondrial Enzyme Markers
ACR exposure resulted in a significant diminution in the activity levels of mitochondrial enzymes. While ACR markedly reduced the activity levels of Complex I-III (NADH-cyt C reductase) (52%), CR treatment significantly improved the activity. A significant (38%) increase in the activity levels of complex I-III was evident at a higher concentration of CR Table 3. Likewise, ACR reduced (by
49%) the MTT reduction property of mitochondria which was restored to normalcy by CR. While ACR caused a marked (87%) increase in ADP/ATP ratio, CR treatment restored the ratio to near normalcy Table 3.
Protective Effect of Crocin supplements on the activity levels of mitochondrial enzyme and ADP/ATP ratio against acrylamide toxicity in C. elegans.
| Groups | NADH-Cytochrome-C reductasea | MTT reducionb | ADP/ATP ratioc | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| CTR | 37.62±1.92 | 4.46±0.19 | 0.46±0.07 | |||||||
| CR (10µM) | 39.92±2.33# | 5.13±0.17* | 0.37±0.04* | |||||||
| ACR (250mg/L) | 18.08±1.55* | 2.3±0.07* | 3.99±0.24* | |||||||
| CR(5 µM)+ACR | 20.86±0.81*# | 2.83±0.09*# | 2.67±0.15*# | |||||||
| CR(10 µM)+ACR | 29.09±0.34*# | 3.03±0.06*# | 2.07±0.06*# |
Table 3: Values are mean ± SE. Data analyzed by one-way ANOVA followed by Tukey’s test (p < 0.05); *= significantly different com
Protective Efficacy of CR on AChE and DA Activity
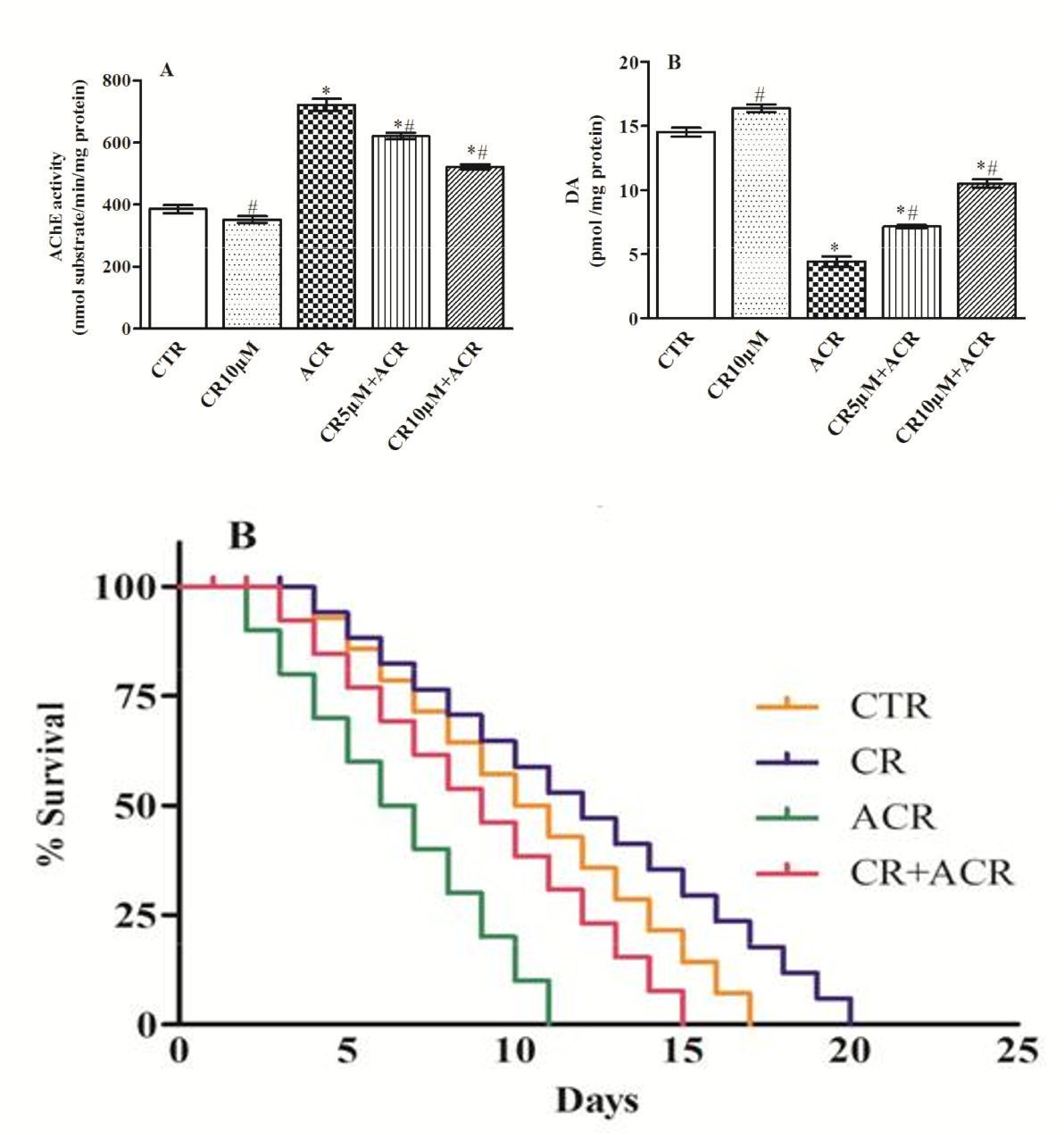
Worms exposed to ACR exhibited a significant (47%) increase in AChE activity compared to controls, while CR treatment diminished (14-28%) the activity levels. Further, the DA content was robustly depleted (69%) among worms exposed to ACR, and CR treatment markedly normalized the levels Figures 4A & B.
CR Treatment Extends Lifespan of N2 and Mutant MEV Strains
Worms exposed to only ACR showed a significant (54%) reduction in survival rate, while worms treated with ACR along with CR showed a marked extension of survival period i.e., 8 days life extension (40%). CR alone treatment could also improve the life span of worms by expanding the lifespan 2 d additional compared to control worms Figure 5A. In another study, well-known short- lived mutant worms (MEV-1) exposed to ACR lived only for 11 d compared to control worms which lived for 17 d. However, CR treatment resulted in marked (36%) extension of life expectancy among these worms Figure 5B. Interestingly, CR alone extended the life of mutant worms significantly (3d more) in comparison to control worms Figure 5B.
Figures 5A & B: Survival curves of worms in different treatment groups: Lifespan of wild type N2 worms (A) and mutant MEV-1 worms (B). Values are percent (%) survival (3 replicates). Data analyzed by Kaplan-Meier (P≤0.0070) followed by Log-rank (Mantel-Cox) Test.
Protective Efficacy of CR To Abrogate ACR Toxicity in BZ555 Strain
When compared to control worms, those treated with ACR exhibited a statistically significant (38%) reduction in the green pixel number. Worms exposed to ACR along with CR exhibited a marked increase in GFP expression (29%) compared to ACR-exposed worms Figure 6.
Discussion
Chronic exposure of children and adult humans to ACR entails understanding the underlying mechanism/s and developing newer strategies to alleviate the adverse effects [50]. While experimental evidences have implicated reproductive toxicity and carcinogenicity owing to ACR exposure, neurotoxicity is the only outcome identified by epidemiological studies among occupationally exposed human populations [17]. Numerous evidences have accumulated over the years which show that ACR induces ataxia, distal muscle weakness and neuronal damage [51, 52]. In view of this, several newer strategies have been attempted to mitigate ACR–mediated neurotoxicity employing experimental models such as Drosophila and rodents [12, 53, 54]. In the present study, we assessed the propensity of CR, a well known carotenoid to modulate ACR neurotoxicity in C. elegans. Saffron and its active constituent CR have been reported to possess various beneficial effects on the central nervous system such as antidepressant, antioxidant, anti-inflammatory, neuromodulant, anticonvulsant, memory enhancing in various studies [29, 55, 56, 57, 58, 59, 60]. Recently we demonstrated the protective effect of CR against rotenone induced neurotoxicity in Drosophila model [29].
Previously few researchers have reported the adverse effects of ACR in C. elegans system. ACR exposure is known to produce adverse effects on metabolic processes, growth, life span and locomotion in worms [21, 61]. Consistent with this, in the present study, worms exposed to ACR showed a significant decrease in egg laying and brood size while CR treatment significantly restored these effects. ACR is reported to be a reproductive toxin affecting male and female reproductive system in rodents [15, 62]. In the present model, ACR exposure significantly decreased the reproductive capability of worms. Interestingly, CR treated worms showed improved reproductive performance clearly suggesting its ability to alleviate the same. This finding corroborates earlier data which have demonstrated that CR exerts protective effects on the development of reproductive organs, fertilization and development of the embryo [63, 64].
Several studies have shown that ACR exposure causes specific phenotypic alterations in experimental models. In the present model, we observed a significant decrease in the locomotor behaviour, thrashing and pharyngeal pumping among worms exposed to ACR which were normalized by CR treatment suggesting its potential protective efficacy. Decreased locomotor activity and DA depletion with ACR exposure have been previously reported in both Drosophila and C. elegans [21, 54].
Growing data in experimental models suggest that increased oxidative stress occurs in almost all neurodegenerative diseases. Oxidative stress refers to disparity between the production of free radicals and the ability of the cell to defend against them. Antioxidants are now being looked upon as convincing curatives against neuronal loss, owing to their potency to neutralize free radicals [65]. ACR is known to cause deleterious effects on antioxidant enzyme activities and elevate oxidative stress in animal models [12, 54, 66]. In the worm model, ACR exposure caused a rigorous oxidative insult as evidenced by significantly elevated levels of ROS and HP and CR co- treatment resulted in a marked reduction. ROS and HP are not just endogenous oxidants, but serve as secondary messengers activating different enzyme cascades activating specific biochemical pathways [67]. Hence the considerable protection rendered by CR treatment against ACR-induced locomotor deficits, as well as the oxidative markers, suggests its potential to modulate ACR mediated oxidative damage.
In the present study, ACR exposure caused a significant depletion in the levels of reduced glutathione and total thiols, whereas, CR treatment significantly restored the levels. Glutathione-allied metabolism is key machinery for cellular protection against conditions of oxidative stress. Glutathione participates in detoxification and may scavenge free radicals, diminish peroxides or be conjugated with electrophonic compounds. Thus, glutathione offers the cell with multiple defenses against a variety of toxins [68]. In the present model, CR caused a significant restoration of ACR-induced activity levels of antioxidant enzymes viz., CAT, GST, TR, and SOD, signifying its efficacy to restore the enzymic antioxidant defense system in worms. Growing number of reports in C. elegans have shown the vital role of antioxidants on enzymic antioxidant defense systems, detoxification of various exogenous peroxides and oxidative stress resistance and their efficacy in life extension [69, 70, 71]. Hence we speculate that the enhanced activity levels of antioxidant enzymes (TR and GST) may be partly responsible for the neuroprotective effect of CR in the current model. Interestingly, we have previously observed a similar effect in Drosophila model with CR supplementation [29]. It would be interesting to examine if CR causes a similar up regulation of antioxidant defences in brain regions under neurotoxicant exposure in rodent models.
Disturbances in neurotransmitter turnover and cholinergic/dopaminergic dysfunctions have been reported amongst the notable features of PD. AChE plays a vital role in the functioning of the neuromuscular system, stopping continuous muscular contraction. Alterations in AChE affect ACh release leading to altered motor coordination. Analysis of the cholinergic function is likely to contribute to further understanding dementia and neurodegenerative diseases [72]. In the present model, ACR exposure resulted in enhanced activity levels of AChE which was brought to near normal range by CR treatment. This finding corroborates earlier reports of AChE inhibitory potency of plant compounds [13, 73]. Dopamine (DA) is a crucial neuromodulator, endowed with diverse roles in the regulation of neural networks controlling sensory modalities, locomotor activity and stereotypical behaviours [74]. In the present study, ACR exposure resulted in significant depletion of DA level consistent with a recent report [21]. Conversely, among ACR flies co-exposed with CR, DA was maintained at basal levels, which is indicative of their prospective specific effect on the dopaminergic neurons. The better performance of worms in the locomotory behaviour may be attributable to the restoration of DA levels among worms co-exposed with CR. Interestingly CR has shown a protective effect in a Drosophila model of Parkinsonism [29].
Besides oxidative stress, mitochondrial dysfunction and decreased ATP production are the key features of aging and neurodegeneration [75]. Recent studies have speculated that disruption or alterations in the activation of pathways including the mitochondrial unfolded protein response (UPRMT) and mitophagy, which regulate mitochondrial function are linked to neurodegenerative diseases due in part to the central role of mitochondria in metabolism, ROS regulation, and proteostasis [76]. In the present model, with ACR exposure we observed significant decrease in the activity levels of complex I-III in worms, which were restored to normalcy with CR treatment. This restorative effect clearly suggests the specific protective effect of CR in modulating the ACR- induced mitochondrial dysfunction. Further, MTT assay was performed to confirm the fate of NADPH-dependant oxidoreductases. Our data revealed the ability of CR to significantly restore the reduced MTT level induced by ACR exposure. The ADP/ATP assay is a quantitative assay revealing the energy status of the cells. ADP/ATP ratio provides a rapid method to measure ADP and ATP levels for the screening of apoptosis, necrosis and proliferation in cells. In the present study, we observed a significant increase in ADP/ATP among ACR exposed worms. Previously, ACR has been shown to affect energy production by inhibiting glycolytic pathway in both in vitro and in vivo models [77, 78]. Reduction in ATPase activity was reported previously illustrating the relationship of ACR-induced neurotoxic targets among rodent and fly models. Alterations in the ADP/ATP ratio have been used to differentiate modes of cell death and viability [12, 13]. While, increased levels of ATP and decreased levels of ADP indicate proliferating cells, conversely, decreased levels of ATP and increased levels of ADP represent apoptotic or necrotic cells [79]. Interestingly, with ACR exposure we evidenced increased levels of ADP/ATP ratio which was effectively attenuated by CR co-treatment.
In the transgenic strain, following ACR exposure we measured neuronal viability by measuring the loss of expression of a GFP in DA neurons. We found a significant loss of GFP expression in CEP dopaminergic neurons after ACR treatment which was visualised by diminished fluorescence in photomicrographs. Interestingly, CR treatment showed a marked increase in GFP expression which is evident by the enhanced increase in green pixel number, a finding which clearly suggests the neuroprotective efficacy of CR in C. elegans. We have previously employed this transgenic strain and demonstrated the potential of Monocrotophos, an organophosphorous insecticide to induce DAergic degeneration and also the modulatory effect of several compounds [80].
Lifespan is a biological process controlled by several genetic and biological factors. Extensive studies on longevity trials in various animal models suggest that the components responsible for lifespan extension affect gene expression, signaling pathways and reduce oxidative stress [81]. Oxidative stress shortens the lifespan of worms. We employed MEV-1 mutant strains which are short lived and more importantly sensitive to oxidative stress and aging. Data obtained in our present lifespan experiment suggests that CR enrichment resulted in a significant extension of lifespan of both wild type N2 worms and also short-lived MEV-1 mutant. MEV-1 mutants which accumulate markers of aging i.e., fluorescent material and protein carbonyls [82, 83]. Further studies are essential to understand the underlying mechanisms by which CR treatment caused increase in life extension.
Conclusion
In summary, our results suggest the propensity of Crocin to alleviate acrylamide-induced phenotype, physiological changes and dopaminergic neurodegeneration in C. elegans. Crocin markedly attenuated ACR induced oxidative stress, mitochondrial dysfunction and significantly enhanced the life span among acrylamide-exposed worms. Taken together, our findings clearly reveal the neurotherapeutic benefits of Crocin and suggest its possible usage as an adjuvant in the management against oxidative –mediated neurodegenerative disorders.
Conflict of Interest
The authors declare that there are no conflicts of interest.
Acknowledgments
The authors thank the Director, CFTRI for his encouragement. The first author thanks the Indian Council of Medical Research (ICMR), New Delhi, for the award of Senior Research Fellowship, while the second author thanks UGC for the award of junior/ Senior Research Fellowships.
References
-
Srivastava R, Ahmed H, Dixit RK, Dharamveer, Saraf SA (2010) _Crocus sativus_ L.: A comprehensive review. Pharmacogn Rev 4(8): 200-208.
-
Ordoudi SA, Befani CD, Nenadis N, Koliakos GG, Tsimidou MZ (2009) Further examination of antiradical properties of _Crocus sativus_ stigmas extract rich in crocins. J Agric Food Chem 57(8): 3080-3086.
-
Selamoglul Z, Ozgen S (2016) Therapeutic Potential of Saffron Crocus (Crocus sativus L.) Turk J Agri Food Sci Tech 4(12): 1240-1245.
-
Purushothuman S (2015) Oxidative stress in neurodegenerative conditions and the protective potential of a natural antioxidant, dietary saffron. Oxid Antioxid Med Sci 4(3): 112-118.
-
Georgiadou G, Grivas V, Tarantilis PA, Pitsikas N (2014) Crocins, the active constituents of Crocus Sativus L., counteracted ketamine–induced behavioural deficits in rats. Psychopharmacology (Berl) 231(4): 717-726.
-
Mehri S, Abnous K, Khooei A, Mousavi SH, Shariaty VM, et al. (2015) Crocin reduced acrylamide-induced neurotoxicity in Wistar rat through inhibition of oxidative stress. Iran J Basic Med Sci 18(9): 902-908.
-
Mohammadzadeh L, Hosseinzadeh H, Abnous K, Razavi BM (2018) Neuroprotective potential of crocin against malathion-induced motor deficit and neurochemical alterations in rats. Environ Sci Pollut Res 25(5): 4904-4914.
-
Heidari S, Mehri S, Hosseinzadeh H (2017) Memory enhancement and protective effects of crocin against D-galactose aging model in the hippocampus of Wistar rats. Iran J Basic Med Sci 20(11): 1250-1259.
-
Razavi BM, Sadeghi M, Abnous K, Hasani FV, Hosseinzadeh H (2017) Study of the role of CREB, BDNF, and VGF neuropeptide in long term antidepressant activity of crocin in the rat cerebellum. Iran J Pharm Res (16): 1452-1462.
-
Smith EA, Oehme FW (2011) Acrylamide and Polyacrylamide: A Review of Production, Use, Environmental Fate and Neurotoxicity. Rev Environ Health 9(4): 215-228.
-
Lee S, Park HR, Lee JY, Cho JH, Song HM, et al. (2018) Learning, memory deficits, and impaired neuronal maturation attributed to acrylamide. J Toxicol Environ Health A 81(9): 254-265.
-
Prasad SN, Muralidhara (2014) Neuroprotective effect of geraniol and curcumin in an acrylamide model of neurotoxicity in _Drosophila melanogaster_: Relevance to neuropathy. J Insect Physiol 60: 7-16.
-
Prasad SN, Muralidhara (2012) Neuroprotective Efficacy of Eugenol and Isoeugenol in Acrylamide- Induced Neuropathy in rats: Behavioral and Biochemical evidence. Neurochem Res 38(2): 330- 345.
-
Santhanasabapathy R, Vasudevan S, Anupriya K, Pabitha R, Sudhandiran G (2015) Farnesol quells oxidative stress, reactive gliosis and inflammation during acrylamide-induced neurotoxicity: Behavioral and biochemical evidence. Neuroscience 308: 212- 227.
-
Wang H, Huang P, Lie T, Li J, Hutz RJ, et al. (2010) Reproductive toxicity of acrylamide-treated male rats. Reprod Toxicol 29(2): 225-230.
-
Xu Y, Cui B, Ran R, Liu Y, Chen H, et al. (2014) Risk assessment, formation, and mitigation of dietary acrylamide: Current status and future prospects. Food Chem Toxicol 69: 1-12.
-
LoPachin RM, (2005) Acrylamide Neurotoxicity: Neurological, Morhological and Molecular Endpoints in Animal Models, In: Friedman M, Mottram D (Eds.), Chemistry and Safety of Acrylamide in Food. Springer US, pp: 21-37.
-
Larguinho M, Costa PM, Sousa G, Costa MH, Diniz MS, et al. (2014) Histopathological findings on Carassius auratus hepatopancreas upon exposure to acrylamide: correlation with genotoxicity and metabolic alterations. J Appl Toxicol 34(12): 1293- 1302.
-
Wang RS , McDaniel LP, Manjanatha MG, Shelton SD, Doerge DR
-
Pennisi M, Malaguarnera G, Puglisi V, Vinciguerra L, Vacante M, et al. (2013) Neurotoxicity of Acrylamide in Exposed Workers. Int J Environ Res Public Health 10(9): 3843-3854.
-
Li J, Li D, Yang Y, Xu T, Li P, et al. (2016) Acrylamide induces locomotor defects and degeneration of dopamine neurons in Caenorhabditis elegans. J Appl Toxicol 36(1): 60-67.
-
Tareke E, Lyn-Cook BD, Duhart H, Newport G, Ali S (2009) Acrylamide decreased dopamine levels and increased 3-nitrotyrosine (3-NT) levels in PC 12 cells. Neurosci Lett 458(2): 89-92.
-
Jayaraj RL, Ravindar DJ, Manigandan K, Padarthi PK, Namasivayam E (2012) An Overview of Parkinson’s Disease and Oxidative Stress: Herbal Scenario. Neuropathol Dis 2(2): 95-122.
-
Chakraborty S, Bornhorst J, Nguyen TT, Aschner M (2013) Oxidative Stress Mechanisms Underlying Parkinson’s Disease-Associated Neurodegeneration in _C. elegans_. Int J Mol Sci 14(11): 23103-23128.
-
O'Reilly LP, Luke CJ, Perlmutter DH, Silverman GA, Pak SC (2014) _C. elegans_ in high-throughput drug discovery. Adv Drug Deliv Rev 69-70: 247-253.
-
Maulik M, Mitra S, Bult-Ito A, Taylor BE, Vayndorf EM (2017) Behavioral Phenotyping and Pathological Indicators of Parkinson’s Disease in _C. elegans_ Models, Front Genet 8:77.
-
Fu RH, Wang YC, Chen CS, Tsai RT, Liu SP, et al. (2014) Acetylcorynoline attenuates dopaminergic neuron degeneration and α-synuclein aggregation in animal models of Parkinson’s disease. Neuropharmacology 82: 108-120.
-
Nidheesh T, Salim C, Rajini PS, Suresh PV (2016) Antioxidant and neuroprotective potential of chitooligomers in _Caenorhabditis elegans_ exposed to Monocrotophos. Carbohydr Polym 135: 138-144.
-
Rao SV, Muralidhara, Yenisetti SC, Rajini PS (2016) Evidence of neuroprotective effects of saffron and crocin in a Drosophila model of Parkinsonism. NeuroToxicology 52: 230-242.
-
Porta-de-la-Riva M, Fontrodona L, Villanueva A, Cerón J (2012) Cerón. Basic _Caenorhabditis_ _elegans_
-
Keaney M, Matthijssens F, Sharpe M, Vanfleteren J, Gems D (2004) Superoxide dismutase mimetics elevate superoxide dismutase activity in vivo but do not retard aging in the nematode _Caenorhabditis_ _elegans_. Free Radic Biol Med 37(2): 239-250.
-
Buckingham SD, Sattelle DB (2009) Fast, automated measurement of nematode swimming (thrashing) without morphometry. BMC Neurosci 10: 84.
-
Hobson RJ, Hapiak VM, Xiao H, Buehrer KL, Komuniecki PR, et al. (2006) SER-7, a _Caenorhabditis_ _elegans_ 5-HT7-like Receptor, Is Essential for the 5-HT Stimulation of Pharyngeal Pumping and Egg Laying. Genetics 172(1): 159-169.
-
Koelle MR, Horvitz HR (1996) EGL-10 Regulates G Protein Signaling in the C. elegans Nervous System and Shares a Conserved Domain with Many Mammalian Proteins. Cell 84(1): 115-125.
-
Bany IA, Dong MQ, Koelle MR (2003) Genetic and cellular basis for acetylcholine inhibition of _Caenorhabditis elegans_ egg-laying behavior. J Neuro sci 23(33): 8060-8069.
-
Middendorf PJ, Dusenbery DB (1993) Fluoroacetic Acid Is a Potent and Specific Inhibitor of Reproduction in the Nematode Caenorhabditis elegans. J Nematol 25(4): 573-577.
-
Chandrashekar KN, Muralidhara (2008) Oxidative alterations induced by d-aspartic acid in prepubertal rat testis in vitro: A mechanistic study. Theriogenology 70(1): 97-104.
-
Wolff SP (1994) Ferrous ion oxidation in the presence of ferric ion indicator xylenol orange for measurement of hydroperoxides. In: Enzymology B-M in (Edns.). Academic Press pp: 182-189.
-
Mokrasch LC, Teschke EJ (1984) Glutathione content of cultured cells and rodent brain regions: A specific fluorometric assay. Anal Biochem 140(2): 506-509.
-
Ellman GL (1959) Tissue sulfhydryl groups, Arch Biochem Biophys 82(1): 70-77.
-
Kostyuk VA, Potapovich AI (1989) Superoxide-- driven oxidation of quercetin and a simple sensitive assay for determination of superoxide dismutase. Biochem Int 19(5): 1117-1124.
-
Aebi H (1984) Catalase in vitro. Methods Enzymol 105: 121-126.
-
Luthman M, Holmgren A (1982) Rat liver thioredoxin and thioredoxin reductase: purification and characterization. Biochemistry (Mosc) 21(26): 6628- 6633.
-
Guthenberg, Alin P, Mannervik B (1985) [64] Glutathione transferase from rat testis, In: Alton Meister (Edns.), Methods in Enzymology, Academic Press 113: 507-510.
-
Ellman GL, Courtney KD, Andres V, Featherstone RM (1961) A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol 7(2): 88-95.
-
Gruntenko NE, Karpova EK, Alekseev AA, Saprykina ZV, Bownes M, et al. (2005) Effects of dopamine on juvenile. hormone metabolism and fitness in Drosophila virilis. J Insect Physiol 51(9): 959-968.
-
Navarro A, Gomez C, López-Cepero JM, Boveris A (2004) Beneficial effects of moderate exercise on mice aging: survival, behavior, oxidative stress, and mitochondrial electron transfer. Am J Physiol - Regul Integr Comp Physiol 286(3): 505-511.
-
Berridge MV, Tan AS (1993) Characterization of the Cellular Reduction of 3-(4,5-dimethylthiazol-2-yl)- 2,5-diphenyltetrazolium bromide (MTT): Subcellular Localization. Substrate Dependence, and Involvement of Mitochondrial Electron Transport in MTT Reduction. Arch Biochem Biophys 303(2): 474-482.
-
Classics Lowry O, Rosebrough N, Farr A, Randall R (1951) Protein measurement with the Folin phenol reagent. J Biol Chem 193: 265-275.
-
Lineback DR, Coughlin JR, Stadler RH, Acrylamide in Foods: A Review of the Science and Future Considerations. Annu Rev Food Sci Technol 3: 15-35.
-
LoPachin RM, Gavin T (2012) Molecular Mechanism of Acrylamide Neurotoxicity: Lessons Learned from Organic Chemistry. Environ Health Perspect 120(12): 1650-1657.
-
LoPachin RM, Gavin T (2008) Acrylamide-Induced Nerve Terminal Damage: Relevance to Neurotoxic and Neurodegenerative Mechanisms. J Agric Food Chem 56(15): 5994-6003.
-
Shinomol GK, Raghunath N, Bharath MM, Muralidhara (2013) Prophylaxis with _Bacopa monnieri_ Attenuates Acrylamide Induced Neurotoxicity and Oxidative Damage via Elevated Antioxidant Function. Cent Nerv Syst Agents Med Chem 13(1): 3-12.
-
Prasad SN, Muralidhara (2012) Evidence of acrylamide induced oxidative stress and neurotoxicity in _Drosophila melanogaster_ – Its amelioration with spice active enrichment: Relevance to neuropathy. NeuroToxicology 33(5): 1254-1264.
-
Amin B, Malekzadeh M, Heidari MR, Hosseinzadeh H (2015) Effect of Crocus sativus extracts and its active constituent safranal on the harmaline-induced tremor in mice. Iran J Basic Med Sci 18(5): 449-458.
-
Papandreou MA, Tsachaki M, Efthimiopoulos S, Cordopatis P, Lamari FN, et al. (2011) Memory enhancing effects of saffron in aged mice are correlated with antioxidant protection. Behav Brain Res 219(2): 197-204.
-
Wang K, Zhang L, Rao W, Su N, Hui H, et al. (2015) Neuroprotective effects of crocin against traumatic brain injury in mice: Involvement of notch signaling pathway. Neurosci Lett 591: 53-58.
-
Wang X, Zhang G, Qiao Y, Feng C, Zhao X (2017) Crocetin attenuates spared nerve injury-induced neuropathic pain in mice. J Pharmacol Sci 135(4): 141-147.
-
Hosseinzadeh H, Talebzadeh F (2005) Anticonvulsant evaluation of safranal and crocin from _Crocus sativus_ in mice. Fitoterapia 76(7-8): 722-724.
-
Ghadrdoost B, Vafaei AA, Rashidy-Pour A, Hajisoltani R, Bandegi AR, et al. (2011) Protective effects of saffron extract and its active constituent crocin against oxidative stress and spatial learning and memory deficits induced by chronic stress in rats. Eur J Pharmacol 667(1-3): 222-229.
-
Hasegawa K, Miwa S, Tsutsumiuchi K, Taniguchi H, Miwa J (2004) Extremely low dose of acrylamide decreases lifespan in Caenorhabditis elegans. Toxicol Lett 152(2): 183-189.
-
Wei Q, Li J, Li X, Zhang L, Shi F (2014) Reproductive toxicity in acrylamide-treated female mice. Reprod Toxicol 46: 121-128.
-
Bakhtiary Z, Shahrooz R, Ahmadi A, Zarei L (2014) Evaluation of antioxidant effects of crocin on sperm quality in cyclophosphamide treated adult mice. Vet Res Forum 5(3): 213-218.
-
Mokhber Maleki E, Eimani H, Bigdeli MR, Ebrahimi B, Shahverdi AH, et al. (2014) A comparative study of saffron aqueous extract and its active ingredient, crocin on the in vitro maturation, in vitro fertilization, and in vitro culture of mouse oocytes. Taiwan J Obstet Gynecol 53(1): 21-25.
-
Shukla V, Mishra SK, Pant HC (2011) Oxidative Stress in Neurodegeneration. Adv Pharmacol Sci 2011: e572634.
-
Yousef MI, El-Demerdash FM (2006) Acrylamide- induced oxidative stress and biochemical perturbations in rats. Toxicology 219(1-3): 133-141.
-
Stone JR, Yang S (2006) Hydrogen Peroxide: A Signaling Messenger. Antioxid Redox Signal 8(3-4): 243-270.
-
Hayes JD, McLellan LI (1999) Glutathione and glutathione-dependent enzymes represent a co- ordinately regulated defence against oxidative stress. Free Radic Res 31(4): 273-300.
-
Ayyadevara S, Engle MR, Singh SP, Dandapat A, Lichti CF, et al. (2005) Lifespan and stress resistance of Caenorhabditis elegans are increased by expression of glutathione transferases capable of metabolizing the lipid peroxidation product 4-hydroxynonenal. Aging Cell 4(5): 257-271.
-
Jee C, Vanoaica L, Lee J, Park BJ, Ahnn J (2005) Thioredoxin is related to life span regulation and oxidative stress response in _Caenorhabditis elegans._ Genes Cells 10(12): 1203-1210.
-
Oláhová M, Taylor SR, Khazaipoul S, Wang J, Morgan BA, et al. (2008) A redox-sensitive peroxiredoxin that is important for longevity has tissue- and stress- specific roles in stress resistance. Proc Natl Acad Sci U S A 105(50): 19839-19844.
-
Craig LA, Hong NS, McDonald RJ (2011) Revisiting the cholinergic hypothesis in the development of Alzheimer’s disease. Neurosci Biobehav Rev 35(6): 1397-1409.
-
Ayaz M, Junaid M, Ahmed J, Ullah F, Sadiq A, et al. (2014) Phenolic contents, antioxidant and anticholinesterase potentials of crude extract, subsequent fractions and crude saponins from _Polygonum hydropiper_ L. BMC Complement Altern Med 14: 145.
-
Zaldivar D, Rauch A, Whittingstall K, Logothetis NK, Goense J (2014) Dopamine-Induced Dissociation of BOLD and Neural Activity in Macaque Visual Cortex. Curr Biol 24 (23): 2805-2811.
-
Lin MT, Beal MF (2006) Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 443(7113): 787-795.
-
Martinez BA, Petersen DA, Gaeta AL, Stanley SP, Caldwell GA, et al. (2017) Dysregulation of the Mitochondrial Unfolded Protein Response Induces Non-Apoptotic Dopaminergic Neurodegeneration in C. elegans Models of Parkinson’s Disease. J Neurosci 37(46): 11085-11100.
-
LoPachin RM, Gavin T (2014) Molecular Mechanisms of Aldehyde Toxicity: A Chemical Perspective. Chem Res Toxicol 27(7): 1081-1091.
-
Martyniuk CJ, Fang B, Koomen JM, Gavin T, Zhang L, et al. (2011) Molecular Mechanism of Glyceraldehyde-3- phosphate Dehydrogenase Inactivation by α,β- Unsaturated Carbonyl Derivatives. Chem Res Toxicol 24(12): 2302-2311.
-
Bradbury DA, Simmons TD, Slater KJ, Crouch SP (2000) Measurement of the ADP: ATP ratio in human leukaemic cell lines can be used as an indicator of cell viability, necrosis and apoptosis. J Immunol Methods 240(1-2): 79-92.
-
Ali SJ, Rajini PS (2012) Elicitation of Dopaminergic Features of Parkinson’s Disease in _C. elegans_ by Monocrotophos, an Organophosphorous Insecticide. CNS Neurol Disord - Drug Targets Former Curr Drug Targets 11(8): 993-1000.
-
Zhu C-T, Ingelmo P, Rand DM (2014) G×G×E for Lifespan in Drosophila: Mitochondrial, Nuclear, and Dietary Interactions that Modify Longevity. PLoS Genet 10(1371): e1004354.
-
Hosokawa H, Ishii N, Ishida H, Ichimori K, Nakazawa H, et al. (1994) Rapid accumulation of fluorescent material with aging in an oxygen-sensitive mutant mev-1 of _Caenorhabditis elegans_. Mech Ageing Dev 74(3): 161-170.
-
Ishii N, Fujii M, Hartman PS, Tsuda M, Yasuda K, et al. (1998) A mutation in succinate dehydrogenase cytochrome b causes oxidative stress and ageing in nematodes. Nature 394(6694): 694-699.
- Hydrogen Peroxide Scavenging by Methanolic Extracts of Coriander: An In Vitro Antioxidant Study
- Aromatherapy in Palliative Care: A Fragrant Quest for Relief
- Empowering Women, Securing Futures: Contraception’s Role in Socioeconomic Progress in India
- Effect of Crospovidone, Croscaramellose Sodium in Combination on the Drug Release of Anti diabetic Medication in Tablet Form
- Knowledge, Attitudes, Anxiety, and Preventive Behaviors Regarding Covid-19 Affliction among Healthcare Workers in Pakistan
- “Competitive Landscape and Brand Equivalents: Implications for ANDA (Abbreviated New Drug Application) Approval”