Detection of Avian Influenza (H5n1) in Some Egyptian Crustaceans from Egypt
To check the liability of some marine and freshwater crustaceans to contract infection with Avian Influenza (H5N1) Randomly 268 different shellfish samples including 28 P. clarckii were collected from Abbassa, Sharkeya city and 240 marine samples (100 P. japonicus, 100 P. semisulcatus, 20 C. sapidus and 20 P. pelagicus) were collected from Suez-Gulf, Suez city and Murrah Lake, Ismailia city. Most of clinically examined Samples were apparently healthy. All marine shellfish samples (Shrimps and Crabs) were negative for H5N1 virus upon using RT-PCR utilizing the highly specific oligonucleotide primers where the 189 bp bands were not detected at any of the tested samples, while only 4/28 Red swamp crayfish were positive. Molecular screening using RT-PCR revealed no infection in all marine examined samples and remarkably showed that 14.4% of freshwater crayfish were positive for H5N1 virus.
Introduction
Despite the fact that, the initial attempts of shrimp farming in Egypt goes back to about 10 years back, the productivity per unit area was never satisfactory due to variety of reasons. The farmed species is considered among those reasons. For years, green tiger shrimp, Penaeus semisulcatus has been tried but with modest productivity. The request to import white leg shrimp, Liptopenaeus vannamei was rejected by fishery authority according to fishery law. Instead, the Indian white prawn was tried starting several years ago with better outcomes [1].
The crab yield, during the period from 2000 to 2015, increased from 19% to 42% of the total lagoon production. Moreover, the results show that the crab catch in Egyptian fisheries is composed mainly of two crab species Portunus pelagicus (L.) and C. sapidus (Rath.). The latter is considered the most dominant in the crab yield with 85% of the total crab production, while C. sapidusis only 15% [2]. Both crab species had migrated to the Egyptian water. Thus, while P. pelagicus (L.) has an Indo-pacific origin and had migrated to the Mediterranean Sea through the Suez-canal, C. sapidus (Rath.) is native to western Atlantic coasts and widely recorded in various Mediterranean regions [3]. The red swamp crayfish, Procambarus clarkii has greatly affected rivers and lakes worldwide [4, 5]. Procambarus clarkii, also known as red swamp crayfish or Louisiana crawfish, is native to northeastern Mexico and the southern USA; it has been introduced in all continents except Antarctica and Oceania, being now considered the most cosmopolitan freshwater crayfish species in the world [6, 7]. In a general way, this species shows good tolerance to a wide range of environmental conditions, elevated adaptive capacity, high growth rate and flexible feeding strategy features that favor its establishment in new available habitats [6, 8]. Avian Influenza is a notifiable pandemic viral disease of poultry and wild birds (migratory and aquatic) caused by an orthomyxovirus Type A. The virus is not adaptable to fish or shellfish cells. However, it can be mechanically lodged on fish mucous, shellfish cuticle or in the intestinal tract or transiently passed to vital immunogenic organs like kidneys of fish or hemolymph of shellfishes [9]. The virus is phylogenetically related to ISA virus of Salmonids and TiLV of Cichlids. Globally, influenza virus has spread from country to country and from continent to another overseas through migratory birds flying pathways (from north to south and vice versa), commercial live poultry export and import and poultry smuggling between different neighbor countries through borders [10, 11]. Influenza viruses were reported to spread from poultry to aquatic animals and vice versa. Marine animals (Whales, Seals, Otters) were reported to have H1, H3, H4, H7 and H13 influenza strains due to predation on infected aquatic birds (Sea gulls, ducks and penguins) (and mechanically or transiently infected aquatic animals (fish, shellfish, bivalves, snails) [11, 12, 13]. H1-2, H3, H4-7, H9-13, H15, H16 Influenza virus strains were reported to spread in wild aquatic environments through predation of fishes on filter feeder animals such as shellfishes (shrimps, lobsters and crayfish), bivalves and snails which might contain the virus for up to 21 days [9, 11]. Such viral strains can also spread through predation of piscivorous birds on shellfishes (shrimps, lobsters and crayfish), bivalves and snails which might contain the virus for up to 21 days followed by virus processing in their gut and consequently releasing it more virulent to poultry populations. They might also be capable of passing the virus with their droppings to settle down into the mucus of the cohabitating fishes which will be further eaten by another aquatic bird giving the virus dynamics an endless epidemiological cycle [9, 11]. Ingestion of droppings of infected aquatic migratory birds is a vulnerable route of spread of these viruses. In fish and shrimp culture, the use of inefficiently treated infected poultry manure in fertilization of earthen pond based facilities can result in spread of certain viral strains e.g. H1-12, H14, H15 within pond water where virus can stay viable in mud and manure for up to 21 days [9]. Virus can be lodged on fish mucus while swimming in poultry manure fertilized earthen ponds and stays for 3 days in mucus till a piscivorous bird (ducks and seagulls) eat it then recirculate the virus after processing in their gut [11]. Further, the filter-feeder shellfishes e.g. shrimp/ crayfish can siphon the virus from the pond bottom where virus can be lodged within haemocytes at their haemolymph and / or in the intestine till being cleared after several days [9]. Fishes and shellfishes are not yet known as specific host for the virus. Thus, they are considered as mechanical / transient biological carrier of the virus. Once carnivorous fish feed on infected dead bird carcasses, they virus find its way through the digestive tract either to blood and from blood to kidneys (hematopoietic organ) or from gut to be shed with feces into the water. In both cases virus can stay viable for up to 21 days without replication [9]. Fish can feed on a filter- feeder aquatic animal (shellfishes, bivalves and snails) who were capable of engulfing the virus through their filter feeding behavior with consequent capture of the virus by their phagocytic haemocytes or by static effect of the hemolymph [9]. Fish will be capable of digesting infected carrier shellfish releasing the virus into the gut which will find its way either to kidney after passing with blood or to the water with fish droppings [11]. In all cases no signs of influenza have been reported in both fish and shellfishes.
Material and Methods
Sampling Locations
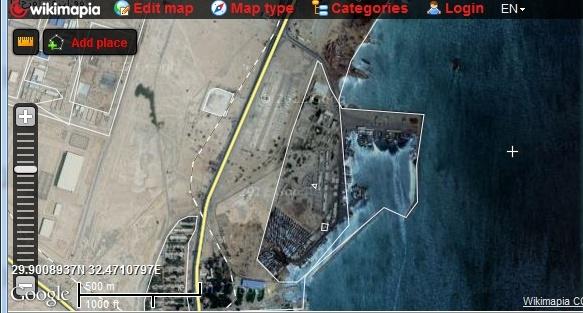
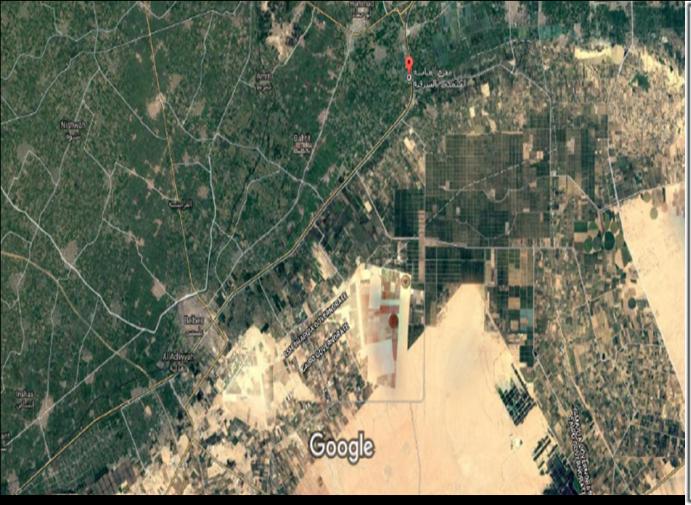
Marine shellfish samples (Shrimp and crabs) were collected from Attaka fishing port, Suez bay at Suez city and Murrah Lake at Ismailia city, Egypt while freshwater crayfish samples were collected from an earthen pond based facilities at Abassa, Sharkiya.

Sampling
Shrimp: A total number of 200 natural adult penalized marine shrimp (50 Penaeus japonicas and 50 Penaeus semisulcatus) were collected from Attaka fishing port, Suez bay at Suez city and the same set of samples were equally collected from Murrah Lake at Ismailia City, Egypt. Crab: A total number of 20 adult Blue Crab (Portugonus pelagicus) were collected from Attaka fishing port, Suez bay at Suez city and 20adult Chesapeake Blue Crab (Canallectus sapidus) were collected from Murrah Lake at Ismailia City, Egypt. Crayfish: A total number of 28 adult Red swamp crayfish (Procambrus clarckii) were collected from earthen pond based aquaculture facilities at Abassa, Sharkiya governorate, Egypt.
Clinical Examination of Collected Shellfishes
Clinical examination was adopted on shrimp, crab and crayfish samples that showed possible disease signs as well as those apparently healthy. Shellfish exoskeleton was cleaned with cotton soaked in 70% ethyl alcohol. Clinical examination of the collected shellfish was done by naked eye for any possible body abnormalities using the method adopted from Lightner and Redman. To apply PM examination on visually inspected shellfish samples, cleaning the surface of the cuticle by cotton soaked in 70% ethyl alcohol then separation of carapace from underlying connective tissue using sterile scissors and forceps were done in clean space or under safety cabinet. Cutting out the Carapace sagittal to expose hepatopancreas for color /consistency and gills, foregut, midgut, hind gut, musculature, periopodes, pleopods, uropod, telson, gonads and genital organs for any deviation in size, color and consistency [14].
Sample Processing
The collected live samples were stored in an insulated ice-box till transferred to the Fish Diseases and Management Laboratory (FDML), Department of Fish Diseases and Management, Faculty of Veterinary Medicine, Cairo University for further processing. Samples were collected during winter and onset of spring and summer seasons of 2016/ 2017 then processed for PCR according to protocols described for marine shrimp / crab samples and for Red swamp crayfish [14]. All collected shellfishes were cleaned, washed three times in sterile distilled water and dried thoroughly with sterile towels before dissection and collection of exoskeleton and internal organs [9]. The carapace of the examined Shrimp / crab / crayfish was split sagittally using a surgical scalpel and scissors. The selected tissues were finely homogenized using sterile homogenizer till consistent homogenates were obtained. Homogenates were further diluted using Hanks balanced salt solution (HBSS: Sigma
Chemical Co, St. Louis, MO, USA) (4 HBSS / 1 homogenate). Haemolymph samples from adult Shrimp/crab/crayfish were collected using sterile syringes then aliquoted into 1 ml microfuge tubes. Diluted homogenates / hemolymph were stored at - 80°C freezer till processed for RNA extraction and PCR detection of Avian Influenza (H5N1).
RNA Extraction
RNA was extracted from the collected samples using Trizol® LS Reagent (Invitrogen, Carlsbad, CA) or the QIAamp® Viral RNA Mini Kit (Qiagen Inc., Valencia, CA) following manufacturers’ instructions with minor modifications. A 0.2 ml sample was homogenized by vortexing and subsequently lysed with 0.4 ml lysis/binding buffer. After binding to the column, DNase-I digestion and washing, the RNA was eluted in 50 μl nuclease-free double-distilled water.
Amplification
Samples were amplified in a one-step RT-PCR in 25 μl final volume, containing 50 mM Tris. HCl pH 8.5, 50 mM NaCl, 7 mM MgCl2, 2 mM DTT, 1 mM each dNTP, 0.4 μM each oligonucleotide, 2.5 U recombinant RNAs in, 10 U AMV reverse transcriptase, 2.5U Ampli-Taq DNA polymerase (all enzymes from Invitrogen) and 5 μl RNA. Primers to detect any type A, influenza viral genome located at M gene as well as the H5-specific primers were adopted from a previous protocol described by [15]. The sequences of the designed primers were MF: 5' CTT CTA ACC GAG GTC GAA ACG 3' and MR: 5' AGG GCA TTT TGG ACA AAG CGT CTA 3') for M gene amplification. The used H5-specific primers were H5F: 5’ ACG TAT GAC TAT TCA CAA TAC TCA G 3’ and H5R: 5’ AGA CCA GCT ACC ATG ATT GC 3’. Thermo-cycling was performed in a thermal cycler (Simpli Amp, Thermos Fisher Scientific, USA- Catalogue # A24811) using the following cycling conditions: 30 minute at 42ºC, 4 minutes at 95ºC once; and 1 minute at 95ºC, 1 minute at 45ºC, 3 minutes at 72ºC repeated 40 times.
Gel Electrophoresis
In two separate 0.1 ml microfuge tubes, five microliters of PCR product in 5μl of 1X TAE Buffer and 1 μl (6X bromophenol) loading dye and 0.5μl of DNA Ladder in 9.5μl of 1X TAE Buffer plus 1 μl loading dye were consistently mixed then 10μl of from each tube were loaded into a 2% agarose gel with ethidium bromide and electrophoresed at 100 V for 30-32 minutes. The resultant bands were viewed using UV Trans-illuminator (Spectronics Corporation, NY, and USA - Catalogue #TVC- 312R). Gel pictures were documented using a digital camera (RX100 V The premium 1.0 type sensor compact camera with superior AF performance, Saint Diego, CA, USA).
Results
Clinical Examination
Shrimp: Clinically, most of the examined shrimp (P. semisulcatus or P. Japonicus) from both Suez Gulf and Murrah Lake were apparently healthy. Crabs: All examined crabs collected from both Suez Gulf and Murrah Lake were apparently healthy with no record of lesions or parasites either externally or internally. Crayfish: Almost all examined crayfish samples from Abassa, Sharkiya were apparently healthy.
Molecular Detection
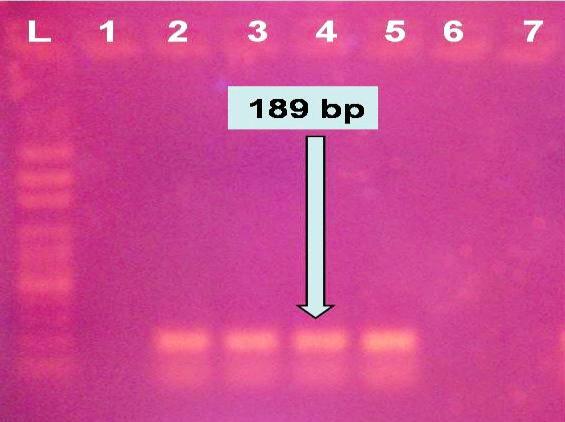
All marine shellfish samples (shrimps and crabs) were negative for H5N1 virus upon using RT-PCR utilizing the highly specific oligonucleotide primers where the 189 bp bands were not detected at any of the tested samples. Only 4 out of 28 Red swamp crayfish were positive for the H5N1 virus documented by detection of 189 bp band.
Total prevalence of AI (H5N1) infection among different examined species
Molecular detection of the H5N1 influenza virus in total of 268 shellfish samples of different marine shrimp (P. japonicas and P. semisulicatus) and crab (Port. pelagicus and C. sapidus) species from both Suez gulf and Murrah Lake revealed negative results (0 out of 40) while the same PCR test revealed that 14.3 % (4 out of 28) of the tested freshwater crayfish (Procambrus clarkii) from Abassa, Sharkiya were positive for the virus as shown in Table 1 and Figure 4. remarkably, all tested pooled tissue samples were positive for the virus with exception of crayfish flesh.
- Geographical
- Type of
- Shellfish
- Prevalence AI
- Positive
- Location (H5N1)
- Results
- P. japonicus
- 0%
- 0 / 50
- P. semisulicatus
- 0%
- 0 / 50
- Port. pelagicus
- 0%
- 0 /20
- Suez gulf
- P. japonicus
- 0%
- 0 / 50
- P. semisulicatus
- 0%
- 0 / 50
- C. sapidus
- 0%
- 0 / 20
- Abassa ,
- Sharkiya
- Murrah Lake
- Procambrus clarckii
- 14.30%
- 4 / 28
Table 1: Total prevalence of AI (H5N1) infection among
Discussion
Despite the fact that Influenza viruses are not adaptable to fish or shellfish cells. However, they can be mechanically lodged on fish mucous, shellfish cuticle or in the intestinal tract or transiently passed to vital immunogenic organs like kidneys of fish or hemolymph of shellfishes being phagocytosed by tissue fixed phagocytes or circulating macrophages [9]. Interestingly, most of orthomyxoviruses affecting fishes are active at freshwater environment e.g. Tilapia lake virus (TiLv) which affect Tilapia species in freshwater lakes and earthen ponds during summer season. However, another Orthomyxovirus like ISA mainly affects Salmonid species at seawater stage while carrier stage is linked to freshwater [11]. In the current study, H5N1 virus was not detected at any of the marine shrimp and crabs collected from either Suez Gulf or Murrah Lake, which contradict the aforementioned findings for other orthomyxoviruses affecting aquatic species like ISA and coincide with TiLV, an emerging orthomyxovirus of the freshwater Tilapias. This, could be attributed to the high salinity levels at both Suez Gulf (35-39 ppt ) and Murrah Lake (41-44 ppt) which has inactivating effect on the virus infectivity as emphasized that avian influenza viruses could be inactivated at salinity levels higher 25 ppt [16, 17, 18]. Interestingly, Red swamp crayfish (P. calrkii) is a bottom filter feeding shellfish which sweeps the pond’s bottom predating on several benthic organisms [19, 20].
Thus, crayfish can engulf bottom settled benthic organism which could be parts of the settled down poultry droppings infected with H5N1 virus or any other pathogens. The well documented fact entailing the accidental presence of some pathogenic viruses (e.g. parvo virus) in the hemolymph of crayfish could explain how H5N1 influenza viral particles were detected in the hemolymph, exoskeleton and internal organs of the 14.3% of the Red swamp crayfish collected from the earthen ponds at Abassa, Sharkiya [21]. The presence of water bodies of Sharkiya province at the scope of major migratory bird flyways, crowd of poultry farms as well as the faulty usage of poultry manure as fish pond fertilizers might explain the possible existence of H5N1 virus in both water and pond’s bottom for enough period that could approach several weeks (up to 190 days in wild viral strains) before being uptake by the filter feeder crayfish [22, 23]. Our results were completely consistent with who reported the presence of H5N1 virus in hemolymph and crusts of Red swamp crayfish with the exception of their flesh which were totally negative for the virus [9]. In conclusion, the current study sheds the light on the magnitude of Influenza virus (H5N1) among wild shrimp, crabs and crayfish populations within the Egyptian open waters. Specifically, we declared that Suez Gulf and Murrah Lake shrimps and crabs populations were refractory to avian influenza (H5N1) infection. Presence of positive freshwater crayfish samples for Influenza (H5N1) confirms the critical epidemiological importance of such invasive aquatic species in spread and establishment of emerging viruses across the Egyptian aquatic habitats as well as their critical significance as a bridge in the Influenza cycle among wild aquatic bird’s populations. Ultimately, results confirm previous assumptions of inability of avian influenza virus (H5N1) to survive higher salinities exceeding 30 ppt.
References
-
Sadek S, Rafael R, Shakouri M, Rafomanana G, Ribeiro FL, et al. (2002) Shrimp aquaculture in Africa and the Middle East: the current reality and trends for the future. Report prepared under the World Bank, NACA, WWF and FAO Consortium Program on Shrimp Farming and the Environment 1-42.
-
Razek FAA, Ismaiel M, Ameran MAA (2016) Occurrence of the blue crab Callinectes sapidus, Rathbun, 1896, and its fisheries biology in Bardawil Lagoon, Sinai Peninsula, Egypt. The Egyptian Journal of Aquatic Research 42(2): 223-229.
-
Lucrezia C, Gianfranco P, Tommaso S, Adele F, Raffaele A (2015) First record of ovegerous Callinectes sapidus (Rathbun, 1896): in the Gargano Lagoons (South-west Adriatic Sea). Bio Invasions Rec 4(4): 281-287.
-
Nalepa TF, Schloesser DW (1993) Zebra Mussels: Biology, Impacts, and Control. CRC Press, Boca Raton, Florida. Office of Technology Assessment. (1993): Harmful Non indigenous Species in the United States, OTA-F-565. United States Government Printing Office, Washington DC.
-
D’Itri FM (1997) Zebra Mussels and Aquatic Nuisance Species. Ann Arbor, University of Michigan Press, 648p.
-
Gherardi F (2006) Crayfish invading Europe: the case study of Procambarus clarkii. Marine and Freshwater Behaviour and Physiology 39(3): 175-191.
-
Chucholl C (2011) Population ecology of an alien “warm water” crayfish (Procambarus clarkii) in a new cold habitat. Knowledge and Management of Aquatic Ecosystems (401): 29.
-
Alcorlo P, Geiger W, Otero M (2004) Feeding preferences and food selection of the red swamp crayfish, Procambarus clarkii, in habitats differing in food item diversity. Crustaceana 77(4): 435-453.
-
Eissa AE, Hussein HA, Zaki MM (2012) Detection of avian influenza (H5N1) in some fish and shellfish from different aquatic habitats across some Egyptian provinces. Life Science Journal 9 (3): 2702-2712.
-
Furuse Y, Suzuki A, Oshitani H (2010) Reassortment between swine influenza A viruses increased their adaptation to humans in pandemic H1N1/09. Infection, Genetics and Evolution 10(4): 569 -574.
-
Eissa AE, Hosni MM (2015) Selected topics in Viral Diseases of Fish and Shellfish. Lap Lambert Publishing 89.
-
Scholtissek C, Naylor E (1988) Fish farming and influenza pandemics. Nature 331(6153): 215.
-
Feare CJ, Yasué M (2006) Asymptomatic infection with highly pathogenic avian influenza H5N1 in wild birds: how sound is the evidence. Virology Journal 3(1): 96.
-
Lightner DV, Redman RM (1998) Shrimp diseases and current diagnostic methods. Aquaculture 164 (1): 201-220.
-
Saberfar E, Forghani-Fard MM, Mosavi M (2007) Multiplex reverse transcriptase-PCR assay for typing and subtyping of influenza A (H5 & H9) virus in Iran. Iranian Biomedical Journal 11(2): 69-74.
-
El-Baky AAA (2012) Studies En Vibriosis In Marine Shrimp Under Egyptian Environment. Master Thesis, Faculty of Veterinary Medicine, Cairo University, Egypt 119.
-
El-Serehy HA, Bahgat MM, Al-Rasheid K, Al-Misned F, Mortuza G, et al. (2014) Cilioprotists as biological indicators for estimating the efficiency of using Gravel Bed Hydroponics System in domestic wastewater treatment. Saudi journal of biological sciences 21(3): 250-255.
-
Brown JD, Goekjian G, Poulson R, Valeika S, Stallknecht DE (2009) Avian influenza virus in water: infectivity is dependent on pH, salinity and temperature. Veterinary microbiology 136(1-2): 20- 26.
-
Momot WT, Gowing H, Jones PD (1978) The dynamics of crayfish and their role in ecosystems. American Midland Naturalist 99(1): 10-35.
-
Scott JR, Thune RL (1986) Ectocommensal protozoan infestations of gills of red swamp crawfish, Procambarus clarkii (Girard), from commercial ponds. Aquaculture 55(3): 161-164.
-
Edgerton B, Owens L (1997) Age at first infection of Cherax quadricarinatus by Cherax quadricarinatus bacilliform virus and Cherax Giardia virus-like virus, and production of putative virus-free crayfish. Aquaculture 152(1-4): 1-12.
-
Melville DS, Shortridge KF (2006) Migratory waterbirds and avian influenza in the East Asian- Australasian Flyway with particular reference to the 2003-2004 H5N1 outbreak. Waterbirds around the world 432-438.
-
Stallknecht DE, Brown JD (2007) Wild birds and the epidemiology of avian influenza. Journal of wildlife diseases 43(3 Supplement): S15-S20.
- hMPV: Is It Another Covid-19 Like Situation?
- Streptomyces: Sources of Novel Discoveries in Antibiotic Research to Combat Antimicrobial Resistance
- A Review of Mosquitoes (Diptera: Culicidae) and Their Biodiversity, Medical and Veterinary Importance
- Past and Current Immunotherapy in Cancer
- Hematological Cancer and Viral Infection
- The Growing Threat of Antimicrobial Resistance in India: Challenges and Solutions