High Mobility Group Box 1 (HMGB1) in Cutaneous Inflammation: An Immune Modulator Bridging Cellular Stress, Ferroptosis and Danger Signaling
High mobility group box 1 (HMGB1) is a dynamic, multifunctional non-histone chromatin protein that translocates between the nucleus, cytoplasm, and extracellular space in response to various stressors. Beyond its nuclear role in chromatin organization and DNA repair, HMGB1 functions as a danger-associated molecular pattern (DAMP), orchestrating autophagy, inflammation, and immune responses through interactions with receptors such as Toll-like receptor 4 (TLR4), the receptor for advanced glycation end-products (RAGE), and CXCR4. This review summarizes HMGB1's structural features, posttranslational modifications (PTMs), and context-dependent signaling across subcellular compartments. Emphasis is placed on its involvement in dermatological conditions including psoriasis, atopic dermatitis, UVB-induced ferroptosis, pigmentation, and photoaging. We further discuss therapeutic modulation of HMGB1, including redox-sensitive interventions and secretiontargeted strategies. Natural compounds such as glycyrrhizin, epigallocatechin gallate (EGCG), and resveratrol demonstrate regulatory effects on HMGB1 activity, supporting their potential roles in skin health and cosmeceutical formulations. A comprehensive understanding of HMGB1’s spatial dynamics and regulatory mechanisms offers novel insights for managing inflammation, barrier dysfunction, and aging in dermatology.
Abbreviations
HMGB1: High Mobility Group Box 1; DAMP: Danger- Associated Molecular Pattern; DNA: Deoxyribonucleic Acid;
PTMs: Post-Translational Modifications; SASP: Senescence- Associated Secretory Phenotype; NLSs: Nuclear Localization Signals.
Introduction
HMGB1 is an evolutionarily conserved, non-histone chromatin protein that was first characterized based on its rapid electrophoretic mobility [1, 2, 3]. Ubiquitously expressed in mammalian cells, HMGB1 serves a fundamental role in nuclear homeostasis by regulating chromatin structure and gene expression [4, 5]. Its structure comprises DNA-binding domains (Box A and Box B) and an acidic C-terminal tail, which allow it to bind and bend DNA, modulate nucleosome stability, and facilitate transcription, replication, and DNA repair. These architectural functions enable HMGB1 to act as a dynamic regulator of genomic organization, essential for cellular integrity and survival [6, 7, 8, 9].
In addition to its nuclear role, HMGB1 exhibits tightly regulated subcellular mobility that responds to various stimuli, including UV radiation, oxidative stress, microbial products, and tissue injury. Post-translational modifications (PTMs) notably acetylation, phosphorylation, methylation, and redox alterations mediate its translocation to the cytoplasm, where it participates in autophagy regulation and inflammatory signaling [10, 11]. In this compartment, HMGB1 participates in the regulation of autophagy and the orchestration of inflammatory signaling. Furthermore, HMGB1 can be secreted extracellularly via active lysosomal pathways or released passively during necrosis and regulated cell death processes such as ferroptosis [12, 13, 14]. In the extracellular environment, HMGB1 functions as a canonical DAMP by engaging PRRs like TLR4, RAGE, and CXCR4, thereby promoting immune cell recruitment, cytokine production, and tissue remodelling [15, 16, 17, 18].
In recent years, HMGB1 has garnered increasing attention in dermatological research due to its multifaceted roles in skin homeostasis and disease [19, 20, 21, 22]. Recent studies have implicated HMGB1 in various dermatological conditions characterized by chronic inflammation, disrupted barrier function, pigmentation disorders, and premature aging [23, 24]. Notably, HMGB1 amplifies UVB-triggered ferroptosis by promoting lipid peroxidation and suppressing glutathione peroxidase 4 (GPX4) activity, thereby exacerbating inflammatory signaling in conditions such as solar dermatitis and psoriasis-like dermatoses. As a redox- sensitive DAMP, HMGB1 acts across nuclear, cytoplasmic, and extracellular compartments to integrate signals from oxidative stress, immune activation, and cellular senescence. Its release is regulated by post-translational modifications such as acetylation and SIRT1-dependent deacetylation, highlighting the importance of epigenetic control [25, 26]. Understanding the spatiotemporal dynamics and regulatory mechanisms governing HMGB1 localization is pivotal for developing targeted interventions that address skin inflammation, barrier dysfunction, and aging.
Recent studies have implicated HMGB1 in various dermatological conditions characterized by chronic inflammation, disrupted barrier function, pigmentation disorders, and premature aging. In psoriasis and atopic dermatitis, extracellular HMGB1 contributes to persistent immune activation and epidermal remodelling [27, 28]. In photoaged skin, UVB-induced oxidative stress promotes HMGB1 translocation and release, which in turn amplifies senescence-associated secretory phenotype (SASP) signaling and matrix degradation [29, 30]. Additionally, HMGB1 modulates melanocyte activity through RAGE-mediated signaling, linking inflammation with hyperpigmentation [31]. As a redox-sensitive molecule, its biological function is finely tuned by its oxidative state, which dictates whether it acts as a cytokine inducer or chemotactic agent [3, 32]. This spatial and redox-dependent plasticity underscores HMGB1’s central role as a molecular switch in cutaneous homeostasis and pathology.
This review provides an integrated overview of HMGB1’s compartment-specific functions, molecular regulation, and translational potential, emphasizing its emerging roles as both a biomarker of inflammatory skin disorders and a therapeutic or cosmeceutical target.
Mechanisms of HMGB1 Translocation and Release
HMGB1’s ability to shuttle between cellular compartments is key to its diverse biological activities. Under basal conditions, HMGB1 predominantly resides in the nucleus, where it contributes to chromatin stability and DNA processing [33]. However, exposure to cellular stress or inflammatory stimuli initiates a tightly orchestrated process of HMGB1 translocation. This is governed by specific post-translational modifications (PTMs) that disrupt its nuclear localization signals (NLSs), facilitating export to the cytoplasm and subsequent secretion [34].
Structural Domains and Binding Features of HMGB1
HMGB1 is encoded by a gene located on chromosome 13q12 and translates into a 215-amino acid polypeptide comprising three key domains: Box A, Box B, and a C-terminal acidic tail [35, 36, 37, 38]. Box A and Box B are both DNA- binding domains, with Box B primarily associated with pro- inflammatory signaling and Box A exhibiting antagonistic, anti-inflammatory properties [36, 39, 40]. These domains form a conserved L-shaped structure with flexible loops, allowing HMGB1 to bind to the minor groove of DNA, bend its helix, and facilitate chromatin decompaction [5]. The acidic tail, rich in negatively charged residues, modulates interactions with histones and nuclear proteins through charge-dependent mechanisms [36, 39, 40].
This domain structure supports HMGB1’s core functions in nucleosome stabilization, chromatin remodelling, and transcriptional regulation [41, 42]. Its conservation across species from rodents to humans underscores the evolutionary importance of these structural motifs [2, 35, 36, 37, 38]. Notably, HMGB1’s dual DNA-binding capacity enables it to act as a context-dependent chromatin organizer or transcriptional co-regulator, with its function modulated by redox state [41, 42]. This structural and functional flexibility positions HMGB1 as a central integrator of nuclear architecture and cellular stress responses.
Post-Translational Modifications (PTMs) of HMGB1
The functional complexity of HMGB1 is significantly expanded through a variety of PTMs, which regulate its localization, protein–protein interactions. Acetylation of lysine residues within the nuclear localization sequences (NLSs) disrupts nuclear retention, and facilitates HMGB1 translocation to the cytoplasm [43, 44]. This process is promoted by histone acetyltransferases (e.g., KAT2B, CBP/p300) and inhibited by deacetylases (e.g., HDAC4) [45, 46, 47], with the TLR4/JAK/STAT1 axis influencing this acetylation balance. [47, 48].
Arginine methylation by PRMTs, such as SETD6, reduces HMGB1’s affinity for TLR4 and modulates cytokine release, offering immunoregulatory effects in inflammatory conditions [49].
Phosphorylation by kinases like protein kinase C (PKC), reduces HMGB1’s DNA-binding capacity, enhances cytoplasmic localization and enhances pro-apoptotic interactions with (e.g., with Bax) [50]. Notably, AMPK- dependent phosphorylation is linked to HMGB1 release during cuproptosis [51].
Redox modifications at cysteines C23, C45, and C106 define HMGB1’s functional. The fully reduced form activates CXCR4 via CXCL12 binding, promoting chemotaxis and tissue repair. The disulfide-bonded form (C23–C45 bond, reduced C106) activates TLR4, initiating pro-inflammatory signaling. Full oxidation (sulfonation of all three cysteines) renders HMGB1 inactive, preventing immune activation [52, 53, 54, 55]. These redox states act as a switch between inflammatory, reparative, and inert modes. These redox states act as a switch between inflammatory, reparative, and inert modes.
Together, these PTMs serve as molecular switches that guide HMGB1’s transition from nuclear sentinel to extracellular danger signal, with implications for inflammation, immunity, and cell death.
Release Pathways
HMGB1 is released into the extracellular space through two distinct pathways: active secretion and passive release due to necrosis or through stress-induced signaling triggered by ultraviolet (UV) irradiation, infection, or mechanical injury. Each mode of release contributes uniquely to the inflammatory milieu [34, 56].
Active secretion occurs through non-classical pathways, as HMGB1 lacks a signal peptide for ER-Golgi-mediated export [57]. Instead, it is translocated into autophagic vesicles or secretory lysosomes, which subsequently undergo lysosomal exocytosis [34]. Exportin-1 facilitates nuclear export of acetylated HMGB1 into the cytoplasm, a step that is primed by upstream PTMs [58]. This route is prominently observed in LPS-stimulated monocytes, but also occurs in keratinocytes, hepatocytes, and endothelial cells under inflammatory or oxidative stress [59].
Passive release is a hallmark of cell death-associated tissue damage. Necrosis, pyroptosis, and ferroptosis all lead to membrane disruption and HMGB1 diffusion into the extracellular space [60, 61]. During ferroptosis, HMGB1 release is not merely a consequence but also an amplifier promoting lipid peroxidation and sustaining the inflammatory loop [47, 62, 63, 64]. Late-stage apoptosis can also result in HMGB1 release, contingent on the redox state of the microenvironment, which determines whether HMGB1 will act as an immunostimulant or remain inert [65].
Collectively, these release pathways illustrate HMGB1’s dual role as both a sensor and effector of cellular stress. While active secretion represents a tightly regulated and often reversible form of immune communication, passive release during ferroptosis and necroptosis serves as an urgent and irreversible alarmin signal, rapidly linking intracellular injury to systemic inflammatory cascades.
Functional Roles of HMGB1 by Compartment
HMGB1’s diverse biological functions are spatially regulated, with its activity contingent upon its localization. In the nucleus, HMGB1 maintains chromatin architecture and genomic integrity [4, 5, 48, 66, 67]. Upon stress-induced translocation to the cytoplasm, it orchestrates autophagy and modulates inflammation through interactions with stress sensors [68, 69, 70]. When released extracellularly, HMGB1 acts as a quintessential alarmin, activating immune responses and contributing to tissue remodelling [71, 72, 73].
Its dynamic redistribution between compartments integrates cellular stress responses with inflammatory signaling. Acting as a DNA chaperone in the nucleus, a stress response modulator in the cytoplasm, and a potent inflammatory mediator in the extracellular milieu, HMGB1 exemplifies structural plasticity and regulatory complexity. The following subsections detail its compartment-specific functions and underlying mechanisms.
Nuclear HMGB1 – Chromatin, DNA Repair, Transcription
Within the nucleus, HMGB1 is indispensable for maintaining genomic homeostasis. By binding to the minor groove of DNA, HMGB1 induces helical bending, which facilitates the recruitment and activity of chromatin remodelling complexes and transcriptional regulators [74, 75]. It stabilizes nucleosomes by modulating histone– DNA interactions, thereby supporting the plasticity of chromatin architecture essential for replication and transcription [48, 67, 76]. HMGB1 also serves as a sentinel of genomic integrity. It interacts with DNA repair proteins such as XPA and APE1 to enhance nucleotide excision and base excision repair, respectively, particularly in response to genotoxic stressors like UV irradiation or oxidative damage [4, 67]. These interactions are vital for preserving genome stability and preventing mutagenesis. Additionally, HMGB1 influences the transcription of genes involved in stress response and immune regulation, linking chromatin dynamics with gene expression.
Cytosolic HMGB1
Upon stress or injury, HMGB1 translocate from the nucleus to the cytoplasm, a process associated with impaired DNA repair capacity and worsened outcomes in models of ischemic and inflammatory tissue damage [77, 78]. This transition highlights the dual role of HMGB1, not only as a nuclear architect involved in chromatin maintenance and genome stability, but also as a key regulator of cellular adaptation to environmental stress.
Once in the cytoplasm, HMGB1 acts as a dynamic stress sensor, integrating metabolic cues and inflammatory signals to coordinate essential cellular processes [69, 70]. Inflammatory stimuli such as lipopolysaccharide (LPS), UVB irradiation, and oxidative stress promote HMGB1 cytoplasmic accumulation, where it governs autophagy initiation and modulates innate immune responses [11, 79, 80, 81]. These cytoplasmic functions enable cells to restore homeostasis or, under excessive stress, initiate protective or programmed death pathways.
Autophagy
Cytosolic HMGB1 plays a pivotal role in the regulation of autophagy, particularly under stress conditions such as nutrient deprivation, oxidative injury, or infection [82, 83, 84, 85]. A key mechanism involves its interaction with Beclin-1 (BECN1), a critical autophagy initiator. HMGB1 competes with Bcl-2 for Beclin-1 binding, thereby releasing Beclin-1 to promote autophagosome formation [34, 86]. This interaction enhances autophagic flux and promotes cell survival during metabolic and inflammatory stress [83, 87, 88].
HMGB1 also interacts with nucleotide-binding oligomerization domain-containing protein 2 (NOD2), facilitating autophagosome maturation and contributing to pathogens clearance and immune regulation, particularly in myeloid cells [89].
In various cancer models, such as gliomas and colorectal cancer, HMGB1-mediated autophagy contributes to tumor adaptation and resistance to therapy [90, 91]. However, in inflammatory bowel disease (IBD), HMGB1 plays an opposing role by preserving intestinal epithelial integrity through sustained autophagic activity, highlighting its context-dependent duality [92].
Interestingly, autophagy and HMGB1 regulate each other bidirectionally. Autophagic vesicles can encapsulate and export HMGB1 via secretory pathways, especially under prolonged stress. This process is notably active in keratinocytes during psoriasis, linking autophagy to chronic skin inflammation [93].
However, dysregulated or excessive autophagy may shift HMGB1’s role toward promoting autophagy-dependent cell death and DAMP release, thereby fuelling inflammatory loops. Thus, the HMGB1–autophagy axis is context- dependent, functioning as a switch between cellular survival and immune activation [87].
Thus, the HMGB1–autophagy axis represents a critical interface between cellular survival and immunological response, balancing repair with inflammation depending on the cellular and tissue context.
Inflammation
Beyond autophagy, cytosolic HMGB1 plays a central role in orchestrating inflammatory responses by functioning as a molecular sensor of nucleic acids and a co-activator of pattern recognition receptor (PRR) signaling. Upon cellular stress or infection, HMGB1 binds to endogenous or pathogen-derived nucleic acids to form immune-stimulatory complexes. These complexes are recognized by endosomal Toll-like receptors (TLRs) particularly TLR3, TLR7, and TLR9 activating downstream IRF3 and IRF7 pathways and promoting the expression of type I interferons and pro- inflammatory cytokines [94].
In parallel, HMGB1 contributes to cytosolic DNA sensing by interacting with the cyclic GMP-AMP synthase (cGAS), which in turn activates the STING (stimulator of interferon genes) pathway. This cascade further amplifies the transcriptional programs involved in antiviral defence and sterile inflammation [95].
HMGB1 also modulates inflammasome activation. It enhances NLRP3 inflammasome assembly and caspase-1 activation, facilitating the maturation and release of IL-1β and IL-18. Through these actions, HMGB1 integrates stress signals into immune effector functions, positioning it at the crossroads of innate immune amplification [96, 97].
These cytoplasmic functions of HMGB1 highlight its role as a molecular bridge linking intracellular danger recognition with broader immunological responses. By modulating the sensitivity and intensity of nucleic acid-sensing pathways, HMGB1 facilitates the elimination of pathogens, but may also exacerbate inflammation-related tissue injury.
Extracellular HMGB1
Once released into the extracellular space either actively via vesicular transport or passively following cell death HMGB1 functions as a canonical damage-associated molecular pattern (DAMP). It binds to several pattern recognition receptors (PRRs), including RAGE, TLR2, TLR4, and CXCR4, initiating robust immune signaling cascades [72, 73].
Receptor Signaling
Through engagement with TLRs and RAGE, HMGB1 activates multiple signaling pathways including NF-κB, MAPKs (ERK, JNK, p38), IRF3, STAT3, and PI3K/AKT thereby inducing the production of pro-inflammatory cytokines and chemokine [71, 98, 99, 100, 101, 102, 103, 104]. These signals coordinate leukocyte recruitment, enhance endothelial permeability, and drive tissue remodelling. Moreover, HMGB1 amplifies immune activation by forming heterocomplexes with molecules such as LPS, IL-1β, or DNA [9, 105, 106].
Beyond its pro-inflammatory functions, HMGB1 also plays a key role in tissue repair. In particular, the HMGB1– CXCL12 complex engages the chemokine receptor CXCR4 to promote chemotaxis, wound healing, and angiogenesis [107, 108]. This ligand-receptor interaction is especially relevant in the context of tissue regeneration and tumor microenvironment signaling, where HMGB1 functions as both a guide and a modulator of cellular trafficking.
Activity Modulation
The immunological functions of extracellular HMGB1 are finely tuned by its redox state. The disulfide-bonded form (C23–C45 intact, C106 reduced) promotes cytokine release through TLR4 signaling [54, 55, 109]. In contrast, fully reduced HMGB1 binds CXCL12 to activate CXCR4-mediated chemotaxis, while fully oxidized HMGB1, where all three cysteines are sulfonated, is biologically inert [110, 111, 112].
Additionally, HMGB1 activity can be curtailed by enzymatic cleavage (e.g., by thrombin) or scavenged by soluble receptors such as sRAGE [113]. These mechanisms serve as critical checkpoints to resolve inflammation and prevent chronic immune activation.
Altogether, extracellular HMGB1 functions as a context- dependent mediator, switching between pro-inflammatory, chemotactic, or inactive roles depending on its redox configuration and receptor landscape. Its regulation represents a promising therapeutic axis for modulating immune responses in inflammatory and regenerative medicine.
HMGB1 in Dermatological Conditions
As a multifunctional DAMP, HMGB1 plays a central role in dermatological pathophysiology through its dynamic localization and context-dependent signaling [114]. This section examines HMGB1’s involvement in key skin conditions, including psoriasis, atopic dermatitis, pigmentation disorders, and aging, and explores its interactions with senescence and ferroptosis-linked pathways.
Psoriasis
Psoriasis is characterized by chronic epidermal inflammation, keratinocyte hyperproliferation, and immune cell infiltration features in which HMGB1 is heavily implicated [115, 116]. Elevated HMGB1 levels in psoriatic skin lesions are driven by autophagy-dependent secretion from stressed keratinocytes and fibroblasts [27, 93, 117, 118]. Once extracellular, HMGB1 binds RAGE and TLRs on immune cells, perpetuating cytokine release (e.g., IL-1β, TNF-α) and sustaining Th17-driven inflammation [119, 120, 121, 122]. Notably, HMGB1 secreted by Langerhans cells further amplifies immune cell recruitment, highlighting its multifaceted role in psoriasis maintenance [123, 124].
Atopic Dermatitis
In atopic dermatitis (AD), a condition marked by epidermal barrier dysfunction and allergic inflammation, HMGB1 contributes to disease flares through its pro- inflammatory actions [125, 126, 127]. Serum levels of HMGB1 correlate with AD severity and flare frequency [128]. Topical or oral treatment with HMGB1-modulating botanicals such as Pruni cortex or ferulic acid attenuates disease severity by disrupting HMGB1-dependent NF-κB and TRPV1 signaling [129, 130]. These findings reinforce the therapeutic utility of targeting HMGB1 to modulate inflammatory circuits and enhance skin resilience.
Skin Aging and Pigmentation
In aging skin, HMGB1 participates in senescence- associated secretory phenotype (SASP) activation [20, 131]. UVB irradiation and pollutants stimulate HMGB1 translocation and release, driving dermal matrix degradation via MMPs and cytokines (e.g., IL-6, IL-1β) [132, 133, 134, 135]. The identification of HMGB1 and Lamin B1 as co- biomarkers in actinic keratosis further supports their utility in senescence profiling [136, 137]. Moreover, UVB irradiation induces ferroptosis a regulated form of cell death which further promotes HMGB1 release from keratinocytes [138]. Importantly, antioxidant and anti-ferroptotic agents may limit HMGB1 release and mitigate photoaging.
Crosstalk with SASP, Melanogenesis, UVB and Ferroptosis
HMGB1 has emerged as a key integrator of cellular senescence, melanogenesis, and ferroptosis pathways in skin biology. In UVB-exposed skin, ferroptotic keratinocytes characterized by lipid peroxidation and glutathione peroxidase 4 (GPX4) suppression release HMGB1 through membrane rupture and exosome-mediated secretion [26, 31, 135, 139]. Extracellular HMGB1 reinforces paracrine inflammatory signaling and promotes melanogenesis by engaging the receptor for advanced glycation end products (RAGE) on melanocytes [19]. This HMGB1–RAGE axis activates PI3K/Akt and MAPK pathways, leading to MITF and tyrosinase upregulation while suppressing pro- apoptotic BAX, thereby supporting melanocyte survival and persistent pigment production [140, 141]. Moreover, HMGB1 establishes a pathological feedback loop in photoaging: sustained oxidative stress enhances its release, which in turn sensitizes keratinocytes to ferroptosis via ACSL4-mediated lipid metabolic reprogramming. Concurrently, HMGB1 primes dermal fibroblasts toward a senescence-associated secretory phenotype (SASP), amplifying chronic inflammation and impairing tissue remodelling [26, 138, 142]. These insights position HMGB1 not merely as a passive alarmin, but as a master regulator of skin homeostasis and dysregulation, with promising potential as a therapeutic and cosmeceutical target.
Therapeutic Modulation of HMGB1
Given HMGB1’s multifaceted roles in inflammatory signaling, cellular stress responses, and tissue remodelling, targeted modulation of its activity presents a compelling avenue for therapeutic intervention in dermatological conditions. This section explores pharmacological and cosmetic strategies that harness natural inhibitors, modulate secretion pathways, or block receptor interactions to mitigate HMGB1-driven pathology.
Natural Inhibitor
Several plant-derived compounds have demonstrated efficacy in downregulating HMGB1 signaling. Glycyrrhizin, a triterpenoid glycoside from liquorice root, directly binds HMGB1 and inhibits its interaction with TLR4 and RAGE [143, 144, 145]. In murine models of atopic dermatitis and psoriasis, glycyrrhizin reduces epidermal HMGB1 levels, suppresses cytokine release, and restores skin barrier integrity [143, 146]. Moreover, glycyrrhizin-mediated HMGB1 blockade alleviates inflammation and pruritus in atopic dermatitis by inhibiting fibroblast activation and disrupting chronic inflammatory circuits [147].
Epigallocatechin gallate (EGCG) also directly binds to HMGB1 and delivers it to lysosomes for degradation in an autophagy-dependent manner in both immune cells and non- immune cells [148]. Notably, EGCG induces aggregation of HMGB1 through large conformational changes and polarized charge redistribution, a process that enhances its lysosomal clearance [149]. Additionally, studies demonstrate that EGCG functions as a TLR4 inhibitor in vivo, attenuating chronic inflammatory responses [150]. Resveratrol inhibits HMGB1 release via an Nrf2-dependent mechanism and downregulates HMGB1-mediated inflammatory cascades by reducing oxidative stress and suppressing NF-κB signaling [151, 152]. Quercetin reduces extracellular HMGB1 while inhibiting downstream inflammatory targets such as RAGE, IL-1β, TNF-α, and COX-2, and simultaneously upregulates Nrf2 to promote cellular antioxidant defence [153]. Senkyunolide I exert anti-inflammatory effects in solar dermatitis by inhibiting ferroptosis-mediated HMGB1 release and disrupting the HMGB1–transferrin receptor 1 (TfR1) positive feedback loop [26]. These natural compounds not only inhibit HMGB1 release and receptor engagement but also modulate its redox and acetylation states, reducing its pro-inflammatory potential. Their dual antioxidant and anti-inflammatory effects highlight their value in applications for managing skin inflammation, hyperpigmentation, and photoaging.
Targeted Strategies: Secretion Inhibition, Receptor Blockade, Redox Regulation
Targeted pharmacological strategies provide refined control over HMGB1 activity by intervening at critical checkpoints in its release or signaling cascade. One approach focuses on preventing its cytoplasmic accumulation. For instance, the inhibition of nuclear export via XPO1 antagonists or activation of histone deacetylases (HDACs) preserves HMGB1 within the nucleus, thereby impeding its secretion. In
parallel, pharmacological agents such as ethyl pyruvate or salicylates suppress HMGB1 release by mitigating oxidative stress and stabilizing nuclear retention. Receptor-level interventions include the use of sRAGE, which acts as a decoy receptor to neutralize HMGB1, and TLR antagonists that prevent receptor engagement and downstream inflammatory signaling. Additionally, redox-modulating compounds can selectively inactivate the pro-inflammatory disulfide form of HMGB1 while sparing its chemotactic isoform. This redox- targeted specificity allows differential modulation of HMGB1’s functions in sterile inflammation versus tissue repair, offering tailored therapeutic windows based on redox status.
Opportunities in Formulations
Compounds such as metformin, when formulated at 6% concentration, have shown efficacy in lowering HMGB1 and IL-1β levels in wound healing models, thereby accelerating epithelial regeneration [154]. In parallel, antioxidants that stabilize HMGB1’s redox state also offer adjunctive benefits in anti-aging regimens by preventing HMGB1-mediated senescence signaling [133, 155, 156].
Taken together, natural modulators of HMGB1 present a robust toolkit for developing skincare solutions. By integrating receptor blockade, secretion inhibition, and oxidative regulation, HMGB1-targeting therapies align closely with current trends in precision dermatology and functional cosmeceutical design. Figure 1 provides a schematic overview of HMGB1’s structural domains, compartment-specific functions, and their relevance to inflammatory skin disorders. The figure summarizes the stepwise translocation of HMGB1 from the nucleus to the extracellular space under stress conditions, and its subsequent activation of pro-inflammatory signaling pathways via TLRs and RAGE. These cascades contribute to the pathogenesis of psoriasis, atopic dermatitis, pigmentation abnormalities, photoaging, and ferroptosis.
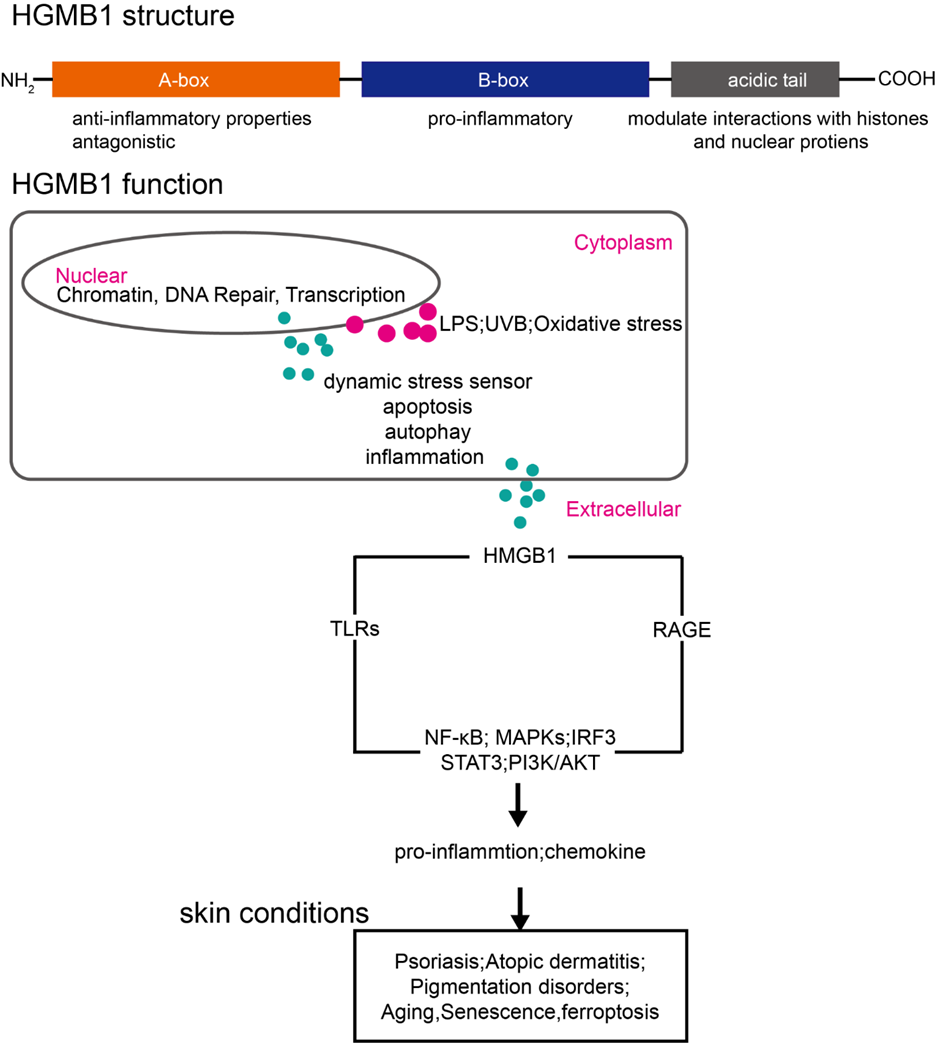
Figure 1: Schematic overview of HMGB1 structure, compartment-specific functions, and relevance in inflammatory skin diseases. HMGB1 consists of three domains: the anti-inflammatory A-box, the pro-inflammatory B-box, and an acidic tail involved in chromatin interactions. In response to stressors (e.g., LPS, UVB, oxidative stimuli), HMGB1 translocates from the nucleus (chromatin regulation) to the cytoplasm (autophagy, inflammation), and ultimately to the extracellular space. There, it activates pattern recognition receptors (TLRs, RAGE), triggering inflammatory pathways (NF-κB, MAPKs, IRF3, STAT3, PI3K/AKT) implicated in skin conditions such as psoriasis, atopic dermatitis, pigmentation disorders, aging, senescence, and ferroptosis.
Perspectives and Future Directions
HMGB1 integrates signals from oxidative stress, immune activation, and cellular senescence, positioning it as a key regulatory node in skin health. Future studies should aim to map HMGB1 functions across different skin cell types and disease states, develop combination strategies using HMGB1- targeting compounds, and explore novel delivery methods to optimize efficacy while minimizing systemic exposure. A nuanced understanding of HMGB1 biology will enable the design of precision-based therapeutic interventions for chronic skin inflammation and aging.
From a therapeutic perspective, development of targeted delivery systems such as liposomal, hydrogel-based, or nanoparticle-formulated inhibitors may improve the bioavailability and precision of HMGB1-targeted compounds while reducing systemic exposure. In parallel, HMGB1’s release patterns, redox isoforms, and receptor interactions offer promising biomarker potential for disease diagnosis, staging, and treatment monitoring in inflammatory and aging-related dermatoses.
Combining HMGB1-targeted strategies with agents that restore barrier function or modulate oxidative stress may offer synergistic benefits. Such multimodal approaches align with the future of personalized dermatological therapeutics and functional cosmetic development.
References
-
Chen R, Kang R, Tang D (2022) The mechanism of HMGB1 secretion and release. Exp Mol Med 54(2): 91–102.
-
Chen R, Zhang Q, Li Y, Wang L, Zhao X, et al. (2013) Emerging role of high mobility group box 1 (HMGB1) in liver diseases. Mol Med 19: 357–366.
-
Sessa L, Bianchi ME (2007) The evolution of High Mobility Group Box (HMGB) chromatin proteins in multicellular animals. Gene 387(1 2): 133–140.
-
Xue J, Sun Y, Chen Z, Fu Y, Wu M, et al. (2021) HMGB1 as a therapeutic target in disease. J Cell Physiol 236(5): 3406–3419.
-
Mandke P, Vasquez KM (2019) Interactions of high mobility group box protein 1 (HMGB1) with nucleic acids: Implications in DNA repair and immune responses. DNA Repair 83: 102701.
-
Stros M (2010) HMGB proteins: interactions with DNA and chromatin. Biochim Biophys Acta Gene Regul Mech 1799(1 2): 101–113.
-
Wang Q, Zhuang S, Zhang Y, Zhang S, Li J, et al. (2007) The HMGB1 acidic tail regulates HMGB1 DNA binding specificity by a unique mechanism. BBRC 360(1): 14–19.
-
Stott K, Watson M, Howe F, Barlow P, Thomas JO, et al. (2010) Tail mediated collapse of HMGB1 is dynamic and occurs via differential binding of the acidic tail to the A and B domains. J Mol Biol 403(5): 706–722.
-
Blair RH, Horn AE, Pazhani Y, Grado L, Goodrich JA, et al. (2016) The HMGB1 C terminal tail regulates DNA bending. J Mol Biol 428(20): 4060–4072.
-
Yang H, Wang H, Czura CJ, Tracey KJ (2013) The many faces of HMGB1: molecular structure functional activity in inflammation, apoptosis, and chemotaxis. J Leukoc Biol 93(6): 865–873.
-
Naglova H, Bucova M (2012) HMGB1 and its physiological and pathological roles. Bratisl Lek Listy 113(3): 163– 171.
-
Wolfson RK, Mapes B, Garcia JG (2014) Excessive mechanical stress increases HMGB1 expression in human lung microvascular endothelial cells via STAT3. Microvasc Res 92: 50–55.
-
El Gazzar M (2007) HMGB1 modulates inflammatory responses in LPS activated macrophages. Inflamm Res 56: 162–167.
-
Bell CW, Jiang W, Reich CF, Pisetsky DS (2006) The extracellular release of HMGB1 during apoptotic cell death. Am J Physiol Cell Physiol 291(6): C1318–1325.
-
Wang H, Yang H, Tracey KJ (2004) Extracellular role of HMGB1 in inflammation and sepsis. J Intern Med 255(3): 320–331.
-
Han J, Zhong Y, Zheng Y, Liu Y, Tan W, et al. (2008) Extracellular high mobility group box 1 acts as an innate immune mediator to enhance autoimmune progression and diabetes onset in NOD mice. Diabetes 57(8): 2118– 2127.
-
Andersson U, Tracey KJ (2011) HMGB1 is a therapeutic target for sterile inflammation and infection. Annu Rev Immunol 29: 139–162.
-
Palumbo R, Galvez BG, Pusterla T, De Marchis F, Cossu G, et al. (2004) Extracellular HMGB1, a signal of tissue damage, induces mesoangioblast migration and proliferation. J Cell Biol 164(3): 441–449.
-
Zhang K, Chen Y, Zhang H, Wang Z, Jin Y, et al. (2019) HMGB1/RAGE mediates UVB induced secretory inflammatory response and resistance to apoptosis in human melanocytes. J Invest Dermatol 139(1): 202–212.
-
Chaichalotornkul S, Jiraungkoorskul W, Anutrakulchai S, Karnjanapiboonwong A, Daduang S, et al. (2015) Secondhand smoke exposure induced nucleocytoplasmic shuttling of HMGB1 in a rat premature skin aging model. BBRC 456(1): 92–97.
-
Satoh TK (2022) The role of HMGB1 in inflammatory skin diseases. J Dermatol Sci 107(2): 58–64.
-
Zhang Q, Li X, Yang J, Wang Y, Chen L, et al. (2012) Role of High Mobility Group Box 1 (HMGB1) in wound healing. J Surg Res 176(1): 343–347.
-
Johnson KE, Wulff BC, Oberyszyn TM, Rossi A, Wilgus TA, et al. (2013) Ultraviolet light exposure stimulates HMGB1 release by keratinocytes. Arch Dermatol Res 305(9): 805–815.
-
Park EJ, Kim YM, Chang KC (2017) Hemin reduces HMGB1 release by UVB in an AMPK/HO-1-dependent pathway in human keratinocytes HaCaT cells. Arch Med Res 48(5): 423–431.
-
Teng Y, Wang Y, Liu H, Li Z, Zhang Y, et al. (2025) Emerging role of ferroptosis in ultraviolet radiation-driven skin photoaging: a narrative review. Photochem Photobiol Sci 24(3): 531–542.
-
Wei Q, Li F, Liu R, Chen H, Zhang J, et al. (2024) Targeting non-classical autophagy-dependent ferroptosis and the subsequent HMGB1/TfR1 feedback loop accounts for alleviating solar dermatitis by senkyunolide I. Free Radic Biol Med 223: 263–280.
-
Chen J, Fu Y, Xiong S (2023) Keratinocyte-derived HMGB1 aggravates psoriasis dermatitis via facilitating inflammatory polarization of macrophages and hyperproliferation of keratinocyte. Mol Immunol 163: 1–12.
-
Dong S, Li D, Shi D (2024) Skin barrier-inflammatory pathway is a driver of the psoriasis atopic dermatitis transition. Front Med 11: 1335551.
-
Chin T, Nguyen T, Yu J, Wang H, Zhao L, et al. (2023) The role of cellular senescence in skin aging and age-related skin pathologies. Front Physiol 14: 1297637.
-
Giroud J, Dubois M, Lemoine A, Vasseur S, Guillon A, et al. (2023) Exploring the communication of the SASP: dynamic, interactive, and adaptive effects on the microenvironment. Int J Mol Sci 24(13): 10788.
-
Wang J, Zhao H, Li W, Xu Q, Deng Y, et al. (2022) The combination of sucrose dilaurate and sucrose laurate suppresses HMGB1: an enhancer of melanocyte dendricity and melanosome transfer to keratinocytes. J Eur Acad Dermatol Venereol 36: 3–11.
-
Andersson U, Tracey KJ, Yang H (2021) Post-translational modification of HMGB1 disulfide bonds in stimulating and inhibiting inflammation. Cells 10(12): 3323.
-
Gupta I, Patel AK (2025) Deciphering HMGB1: Across a spectrum of DNA and nucleosome dynamics. Cell Biol Int 49(3): 235–249.
-
Chen R, Kang R, Tang D (2022) The mechanism of HMGB1 secretion and release. Exp Mol Med 54(2): 91–102.
-
Datta S, Biswas A, Mandal M, Pal S, Roy S, et al. (2023) High Mobility Group Box 1 (HMGB1): Molecular signaling and potential therapeutic strategies. Cells 13(23): 1946.
-
Gong W, Fan H, Liu J, Wu Y, Zhang J, et al. (2010) The anti- inflammatory activity of HMGB1 A box is enhanced when fused with C-terminal acidic tail. J Biomed Biotechnol 2010: 915234.
-
Li J, Wang H, Mason JM, Levine J, Yu M, et al. (2003) Structural basis for the proinflammatory cytokine activity of high mobility group box 1. Mol Med 9(1-2): 37–45.
-
Stros M (2009) HMGB proteins: interactions with DNA and chromatin. Biochim Biophys Acta 1799(1-2): 101– 113.
-
Lotze MT, Tracey KJ (2005) High-mobility group box 1 protein (HMGB1): nuclear weapon in the immune arsenal. Nat Rev Immunol 5(4): 331–342.
-
Li J, Zhao Y, Liu J, Wang C, Zhang Y, et al. (2024) HMGB1: A new target for ischemic stroke and hemorrhagic transformation. Transl Stroke Res.
-
Musumeci D, Roviello GN, Montesarchio D (2014) An overview on HMGB1 inhibitors as potential therapeutic agents in HMGB1-related pathologies. Pharmacol Ther 141(3): 347–357.
-
Chikhirzhina E (2020) Functional diversity of non- histone chromosomal protein HmgB1. Int J Mol Sci 21(21): 7948.
-
Yang Z (2014) PARP-1 mediates LPS-induced HMGB1 release by macrophages through regulation of HMGB1 acetylation. J Immunol 193(12): 6114–6123.
-
Bonaldi T (2003) Monocytic cells hyperacetylate chromatin protein HMGB1 to redirect it towards secretion. EMBO J.
-
Yang K (2020) Lactate promotes macrophage HMGB1 lactylation, acetylation, and exosomal release in polymicrobial sepsis. Cell Death Differ 29(1): 133–146.
-
Wang Y, Wang L, Gong Z (2019) Regulation of acetylation in high mobility group protein B1 cytosol translocation. DNA Cell Biol 38(5): 491–499.
-
Park EJ (2018) Degradation of histone deacetylase 4 via the TLR4/JAK/STAT1 signaling pathway promotes the acetylation of high mobility group box 1 (HMGB1) in lipopolysaccharide activated macrophages. FEBS Open Bio 8(7): 1119–1126.
-
Tang D (2023) The multifunctional protein HMGB1: 50 years of discovery. Nat Rev Immunol 23(12): 824–841.
-
Walter CEJ (2020) Bladder neoplasms and NF κB: an unfathomed association. Expert Rev Mol Diagn 20(5): 497–508.
-
Wang X (2024) Phosphorylation by Protein Kinase C Weakens DNA Binding Affinity and Folding Stability of the HMGB1 Protein. Biochemistry 63(14): 1718–1722.
-
Liu J, Zhang X, Yang Y, You T, Zhang S, et al. (2022) HMGB1 is a mediator of cuproptosis-related sterile inflammation. Front Cell Dev Biol 10: 996307.
-
An K, Wang F, Li G, Chen Q, Chen C, et al. (2022) Spinal PKC activation induced neuronal HMGB1 translocation contributes to hyperalgesia in a bone cancer pain model in rats. Exp Neurol 303: 80–94.
-
He J, Wang L, Zhang R, Li S, Chen X, et al. (2022) Lagopsis supina ameliorates myocardial ischemia injury by regulating angiogenesis, thrombosis, inflammation, and energy metabolism through VEGF, ROS and HMGB1 signaling pathways in rats. Phytomedicine 120: 155050.
-
Janko C (2014) Redox modulation of HMGB1-related signaling. Antioxid Redox Signal 20(7): 1075–1085.
-
Tang D, Kang R, Livesey KM, Zeh HJ, Lotze MT (2011) High-mobility group box 1, oxidative stress, and disease. Antioxid Redox Signal 14(7): 1315–1335.
-
Scaffidi P, Misteli T, Bianchi ME (2002) Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 418(6894): 191–195.
-
Kepes F, Rambourg A, Satiat B (2005) Secretory Pathway. Int Rev Cytol 242: 55.
-
Gonzalez M (2024) HMGB1 expression levels correlate with response to immunotherapy in non-small cell lung cancer. Lung Cancer Targets Ther 55–67.
-
Gardella S (2002) The nuclear protein HMGB1 is secreted by monocytes via a non clasical vesicle mediated secretory pathway. EMBO Rep.
-
Scaffidi P, Misteli T, Bianchi ME (2002) Release of chromatin protein HMGB1 by necrotic cells triggers inflammation. Nature 418(6894): 191–195.
-
Kepes F, Rambourg A, Satiat Jeunemaitre B (2005) Secretory pathway mechanisms of HMGB1 release. Int Rev Cytol 242: 55.
-
Wen Q, Zhang L, Chen S, Li W, Xu Y, et al. (2019) The release and activity of HMGB1 in ferroptosis. Biochem Biophys Res Commun 510(2): 278–283.
-
Ye F, Wang L, Li X, Zhang Y, Chen H, et al. (2019) HMGB1 regulates erastin induced ferroptosis via RAS JNK/p38 signaling in HL 60/NRASQ61L cells. Am J Cancer Res 9(4): 730.
-
Wu Y, Zhang Q, Li H, Zhou J, Xu Z, et al. (2021) HMGB1 regulates ferroptosis through Nrf2 pathway in mesangial cells in response to high glucose. Biosci Rep 41(2): BSR20202924.
-
Bell CW, Jiang W, Reich CF, Pisetsky DS (2006) The extracellular release of HMGB1 during apoptotic cell death. Am J Physiol Cell Physiol 291(6): C1318–1325.
-
Thomas JO, Stott K (2012) H1 and HMGB1: modulators of chromatin structure. Biochem Soc Trans 40(2): 341–346.
-
Stros M (2010) HMGB proteins: interactions with DNA and chromatin. Biochim Biophys Acta Gene Regul Mech 1799(1 2): 101–113.
-
Tang D, Kang R, Livesey KM, Zeh HJ, Lotze MT (2010) Endogenous HMGB1 regulates autophagy. J Cell Biol 190(5): 881–892.
-
Zhu X, Wang Y, Liu J, Chen Z, Yang Q, et al. (2015) Cytosolic HMGB1 controls the cellular autophagy/apoptosis checkpoint during inflammation. J Clin Invest 125(3): 1098–1110.
-
Tang D, Kang R, Zeh HJ, Lotze MT, et al. (2010) HMGB1 release and redox regulates autophagy and apoptosis in cancer cells. Oncogene 29(38): 5299–5310.
-
Sims GP, Rowe DC, Rietdijk ST, Herbst R, Coyle AJ (2010) HMGB1 and RAGE in inflammation and cancer. Annu Rev Immunol 28: 367–388.
-
Harris HE, Andersson U, Pisetsky DS (2012) HMGB1: a multifunctional alarmin driving autoimmune and inflammatory disease. Nat Rev Rheumatol 8(4): 195–202.
-
Ozbay Kurt FG, Smith J, Lee Y, Patel A, Zheng G, et al. (2024) S100A9 and HMGB1 orchestrate MDSC mediated immunosuppression in melanoma through TLR4 signaling. J Immunother Cancer 12(9).
-
McCauley MJ, Johnson RC, Williams KP, Thompson MP, Robertson DL, et al. (2007) HMGB binding to DNA: single and double box motifs. J Mol Biol 374(4): 993–1004.
-
Cato L, Visconti R, DelGallo M, DeMarchis F, Bianchi ME (2008) The interaction of HMGB1 and linker histones occurs through their acidic and basic tails. J Mol Biol 384(5): 1262–1272.
-
Kozlova AL, Fedorov AO, Kirik OV, Petrova MV, Sokolov DV, et al. (2018) HMGB proteins as DNA chaperones that modulate chromatin activity. Mol Biol 52(5): 637–647.
-
Miao J, Wu Y, Chen Z, Zhang H, Li F, et al. (2020) Nuclear HMGB1 promotes the phagocytic ability of macrophages. Exp Cell Res 393(1): 112037.
-
Zhao G, Zhu D, Wang L, He Y, Li Q, et al. (2017) Down regulation of nuclear HMGB1 reduces ischemia induced HMGB1 translocation and release and protects against liver ischemia reperfusion injury. Sci Rep 7(1): 46272.
-
Deng M, Tang Y, Li W, Wang X, Zhang R, et al. (2019) Location is the key to function: HMGB1 in sepsis and trauma induced inflammation. J Leukoc Biol 106(1): 161–169.
-
Stavely R, Nguyen T, Smith J, Lee M, Patel C, et al. (2022) Oxidative stress induced HMGB1 translocation in myenteric neurons contributes to neuropathy in colitis. Biomolecules 12(12): 1831.
-
Mou K, Zhang L, Li Y, Sun P, Wang H, et al. (2017) HMGB1/RAGE axis promotes autophagy and protects keratinocytes from ultraviolet radiation induced cell death. J Dermatol Sci 85(3): 162–169.
-
Kang R, Livesey KM, Zeh HJ, Lotze MT, Tang D (2011) HMGB1 as an autophagy sensor in oxidative stress. Autophagy 7(8): 904–906.
-
Chen R, Kang R, Tang D (2024) HMGB1 in the interplay between autophagy and apoptosis in cancer. Cancer Lett 581: 216494.
-
Chen R, Kang R, Tang D (2022) The mechanism of HMGB1 secretion and release. Exp Mol Med 54(2): 91–102.
-
Devis-Jauregui L, Smith J, Brown K, Davis H, Wilson A, et al. (2021) Autophagy in the physiological endometrium and cancer. Autophagy 17(5): 1077–1095.
-
Kang R, Livesey KM, Zeh HJ, Lotze MT, Tang D (2011) Metabolic regulation by HMGB1 mediated autophagy and mitophagy. Autophagy 7(10): 1256–1258.
-
Wen Q, Zhang L, Chen S, Li W, Xu Y, et al. (2019) The release and activity of HMGB1 in ferroptosis. Biochem Biophys Res Commun 510(2): 278–283.
-
Luo Y, Smith J, Adams M, Johnson K, Wang T, et al. (2014) Cancer usurps skeletal muscle as an energy repository. Cancer Res 74(1): 330–340.
-
Kim SW, Park EJ, Lee SJ, Choi YJ, Seo YK (2022) Cytosolic HMGB1 mediates LPS induced autophagy in microglia by interacting with NOD2 and suppresses its proinflammatory function. Cells 11(15).
-
Zhao M, Wang Y, Li X, Zhou J, Fu Q, Li L, et al. (2021) YAP promotes autophagy and progression of gliomas via upregulating HMGB1. J Exp Clin Cancer Res 40: 1–15.
-
Liu W, Chen X, Tang D (2015) HMGB1-mediated autophagy modulates sensitivity of colorectal cancer cells to oxaliplatin via MEK/ERK signaling pathway. Cancer Biol Ther 16(4): 511–517.
-
Zhu X, Wang Y, Liu J, Chen Z, Yang Q, et al. (2015) Cytosolic HMGB1 controls the cellular autophagy/apoptosis checkpoint during inflammation. J Clin Invest 125(3): 1098–1110.
-
Wang Z, Li Y, Xu J, Zhao Q, Chen G, et al. (2021) Autophagy-based unconventional secretion of HMGB1 by keratinocytes plays a pivotal role in psoriatic skin inflammation. Autophagy 17(2): 529–552.
-
Yanai H, Ban T, Taniguchi T (2011) Essential role of high- mobility group box proteins in nucleic acid-mediated innate immune responses. J Intern Med 270(4): 301– 308.
-
Wang Z, Xu J, Li Q, Zhang Y, Chen G, et al. (2019) cGAS/ STING axis mediates a topoisomerase II inhibitor- induced tumor immunogenicity. J Clin Invest 129(11): 4850–4862.
-
Wang W, Li L, Zheng X, Song Y, Zhang H, et al. (2019) Upregulation of PD L1 via HMGB1-activated IRF3 and NF κB contributes to UV radiation induced immune suppression. Cancer Res 79(11): 2909–2922.
-
Olcum M, Demirel B, Erman F, Yilmaz S, Mete E, et al. (2021) Ethyl pyruvate attenuates microglial NLRP3 inflammasome activation via inhibition of HMGB1/NF κB/miR 223 signaling. Antioxidants (Basel) 10(5).
-
Wang S, Zhang Y (2020) HMGB1 in inflammation and cancer. J Hematol Oncol 13(1): 116.
-
Shao M, Li X, Zhang Y, Chen S, Fan Z (2016) Rac1 mediates HMGB1 induced hyperpermeability in pulmonary microvascular endothelial cells via MAPK signaling. Mol Med Rep 13(1): 529–535.
-
Luo L, Zhang X, Mei Y, Zhang X, Wang Y (2022) Inhibition of inflammatory liver injury by the HMGB1-A box through HMGB1/TLR-4/NF κB signaling in an acute liver failure mouse model. Front Pharmacol 13: 990087.
-
Chen X, Li Y, Zhao Z, Wang W, Zhao X (2018) Omega-3 polyunsaturated fatty acid attenuates the inflammatory response by modulating microglia polarization through SIRT1-mediated deacetylation of the HMGB1/NF κB pathway following experimental traumatic brain injury. J Neuroinflammation 15(1): 116.
-
Chen G, Zhang Z, Yang H, Wang L, Wu J (2021) ALKBH5-modified HMGB1 STING activation contributes to radiation-induced liver disease via innate immune response. Int J Radiat Oncol Biol Phys 111(2): 491–501.
-
Zhang YG, Wu LN, Liu NX, Zhou Y, Li L (2019) Intestinal epithelial HMGB1 inhibits bacterial infection via STAT3 regulation of autophagy. Autophagy 15(11): 1935–1953.
-
Feng Z, Li X, Zhang L, Sun P, Chen Y (2024) Inhibition of ferroptosis rescues M2 macrophages and alleviates arthritis by suppressing the HMGB1/TLR4/STAT3 axis in M1 macrophages. Redox Biol 75: 103255.
-
Ren Y, Li D, Wang X, Liu Q, Zhang J (2023) HMGB1: a double-edged sword and therapeutic target in the female reproductive system. Front Immunol 14: 1238785.
-
Shen P, Liu J, Zhang W, Chen Y, Zhao X (2024) Targeting HMGB1 and its interaction with receptors: challenges and future directions. J Med Chem 67(24): 21671–21694.
-
Pirani E, Rossi L, Bianchi F, Morini M, Santoro N (2024) Tumor cells express and maintain HMGB1 in the reduced isoform to enhance CXCR4-mediated migration. Front Immunol 15: 1358800.
-
Haque N, Singh P, Kumar V, Namdev S, Sharma P (2020) Role of the CXCR4 SDF1 HMGB1 pathway in the directional migration of cells and regeneration of affected organs. World J Stem Cells 12(9): 938–951.
-
Kwak MS, Park SH, Kim HJ, Lee JY, Choi JY (2021) Reactive oxygen species induce Cys106 mediated anti parallel HMGB1 dimerization that protects against DNA damage. Redox Biol 40: 101858.
-
Schiraldi M, Raucci A, Munoz LM, Livoti E, Celona B, et al. (2012) HMGB1 promotes recruitment of inflammatory cells to damaged tissues by forming a complex with CXCL12 and signaling via CXCR4. J Exp Med 209(3): 551–563.
-
Janko C (2014) Redox modulation of HMGB1-related signaling. Antioxidants & redox signaling, 2014. 20(7): 1075–1085.
-
Yang H (2021) Redox modifications of cysteine residues regulate the cytokine activity of HMGB1. Molecular Medicine 27: 1–7.
-
Zhang F (2017) sRAGE alleviates neutrophilic asthma by blocking HMGB1/RAGE signalling in airway dendritic cells. Sci Rep 7(1): 14268.
-
Satoh TK (2022) The role of HMGB1 in inflammatory skin diseases. J Dermatol Sci 107(2): 58–64.
-
Chen T (2013) Increased HMGB1 serum levels and altered HMGB1 expression in patients with psoriasis vulgaris. Archives of dermatological research 305: 263– 267.
-
Zhu X (2024) HMGB1 regulates Th17 cell differentiation and function in patients with psoriasis. Immunity, Inflammation and Disease 12(2): e1205.
-
Lee CY, Jeong KH (2020) 16202 Effect of high- mobility group box 1 on keratinocyte and fibroblast and its involvement in the development of psoriasis. Journal of the American Academy of Dermatology 83(6): AB172.
-
Strohbuecker L (2019) Increased dermal expression of chromatin-associated protein HMGB1 and concomitant T-cell expression of the DNA RAGE in patients with psoriasis vulgaris. Psoriasis: Targets and Therapy 2019: 7–17.
-
Martinotti S, Patrone M, Ranzato E (2015) Emerging roles for HMGB1 protein in immunity, inflammation, and cancer. Immunotargets Ther 4: 101–109.
-
Bianchi ME, Crippa MP, Manfredi AA, Querini P, Venereau E, et al. (2017) High-mobility group box 1 protein orchestrates responses to tissue damage via inflammation, innate and adaptive immunity, and tissue repair. Immunol Rev 280(1): 74–82.
-
Wang Z, Zhao Y, Wang Y, Gao Y, Zhang F, et al. (2021) Autophagy-based unconventional secretion of HMGB1 by keratinocytes plays a pivotal role in psoriatic skin inflammation. Autophagy 17(2): 529–552.
-
Chen J, Fu Y, Xiong S (2023) Keratinocyte derived HMGB1 aggravates psoriasis dermatitis via facilitating inflammatory polarization of macrophages and hyperproliferation of keratinocyte. Mol Immunol 163: 1–12.
-
Yuan J, Li J, Wu Y, Yang J, Tang D (2024) HMGB1 as an extracellular pro-inflammatory cytokine: Implications for drug-induced organic damage. Cell Biol Toxicol 40(1): 55.
-
Gao Y, Zhang X, Yuan H, Wang Z, Fang Y (2024) Damage-Associated Molecular Patterns, a class of potential psoriasis drug targets. Int J Mol Sci 25(2): 771.
-
Karuppagounder V, Arumugam S, Thandavarayan RA, Sreedhar R, Watanabe K, et al. (2015) Modulation of HMGB1 translocation and RAGE/NF-κB cascade by quercetin treatment mitigates atopic dermatitis in NC/ Nga transgenic mice. Exp Dermatol 24(6): 418–423.
-
Bieber T (2022) Atopic dermatitis: an expanding therapeutic pipeline for a complex disease. Nat Rev Drug Discov 21(1): 21–40.
-
Casciaro M, Di Salvo E, Gangemi S (2021) HMGB-1 in Psoriasis. Biomolecules 12(1): 60.
-
Cuppari C, Salpietro A, Romeo C, Berretta M, Corsello G, Salpietro V (2016) HMGB1 levels in children with atopic eczema/dermatitis syndrome (AEDS). Pediatr Allergy Immunol 27(1).
-
Watanabe K, Kageyama-Yahara N, Tsuboi K, Ohta T, Tsuji NM, et al. (2015) Pruni cortex ameliorates skin inflammation possibly through HMGB1-NFκB pathway in house dust mite induced atopic dermatitis NC/Nga transgenic mice. J Clin Biochem Nutr 56(3): 186–194.
-
Wang HM, Chen J, Li X, Zhang L, Xu Q, et al. (2023) Ferulic acid alleviates inflammatory manifestations in atopic dermatitis through modulation of the TRPV1/ HMGB1 signaling pathway. J Dermatol Sci.
-
Zhao J, Li Y, Chen X, Yang Z, Liu S, et al. (2018) Epidermal HMGB1 activates dermal fibroblasts and causes hypertrophic scar formation in reduced hydration. J Invest Dermatol 138(11): 2322–2332.
-
Biran A, Reva A, Krizhanovsky V, Segal E, Amariglio N, et al. (2017) Quantitative identification of senescent cells in aging and disease. Aging Cell 16(4): 661–671.
-
Davalos AR, Coppe JP, Campisi J, Desprez PY (2013) p53 dependent release of Alarmin HMGB1 is a central mediator of senescent phenotypes. J Cell Biol 201(4): 613–629.
-
Sofiadis K, Diamanti E, Papadaki M, Kotsopoulou A, Papadopoulou E, et al. (2021) HMGB1 coordinates SASP related chromatin folding and RNA homeostasis on the path to senescence. Mol Syst Biol 17(6): e9760.
-
Johnson KE, Ferrick DA, Tsai CH, Capicciotti CJ, et al. (2013) Ultraviolet light exposure stimulates HMGB1 release by keratinocytes. Arch Dermatol Res 305: 805– 815.
-
Wang AS, Athar M, Sorg NT, Wilson CL, et al. (2022) Identification and quantification of senescent cell types by lamin B1 and HMGB1 in Actinic keratosis lesions. J Dermatol Sci 105(1): 61–64.
-
Liani O (2019) In Vitro and In Vivo Characterization of High Mobility Group Box 1 (HMGB1) and Lamin B1 (LMNB1) as senescence biomarkers in skin cell types. Indonesia International Institute for Life Sciences.
-
Vats K, Zhang Y, Zahiruddin S, Nielsen JK, et al. (2021) Keratinocyte death by ferroptosis initiates skin inflammation after UVB exposure. Redox Biol 47: 102143.
-
Xiao T, Liu R, Chen Z, Wu Y, et al. (2024) ATG5 mediated keratinocyte ferroptosis promotes M1 polarization of macrophages to aggravate UVB induced skin inflammation. J Photochem Photobiol B 257: 112948.
-
Wang J, Liu Y, Zhang Y, Li J, Liu H, et al. (2022) The combination of sucrose dilaurate and sucrose laurate suppresses HMGB1: an enhancer of melanocyte dendricity and melanosome transfer to keratinocytes. J Eur Acad Dermatol Venereol 36 Suppl 3: 3–11.
-
Zhang L, Gao J, Li Z, Xu Y, Zhao W, et al. (2025) Effectiveness and Safety of Oral Compound Glycyrrhizin Followed by Phototherapy for the Treatment of Progressive Vitiligo in Children. Pigment Cell Melanoma Res 38(2): e13226.
-
Liu L, Kang X (2022) ACSL4 is overexpressed in psoriasis and enhances inflammatory responses by activating ferroptosis. Biochem Biophys Res Commun 623: 1–8.
-
Wang Y, Liu Y, Wang J, Li X, Yu J, et al. (2018) Glycyrrhizin ameliorates atopic dermatitis like symptoms through inhibition of HMGB1. Int Immunopharmacol 60: 9–17.
-
Mollica L, De Marchis F, Spitaleri A, Dallacasa T, et al. (2007) Glycyrrhizin binds to high mobility group box 1 protein and inhibits its cytokine activities. Chem Biol 14(4): 431–444.
-
Vergoten G, Bailly C (2020) Analysis of glycyrrhizin binding to protein HMGB1. Med Drug Discov 7: 100058.
-
Qiong H, Zhang D, Li Y, Gao C, Li M, et al. (2021) Glycyrrhizin improves the pathogenesis of psoriasis partially through IL 17A and the SIRT1 STAT3 axis. BMC Immunol 22(1): 34.
-
Zhou L, Huang Z, Wang Y, Guo X, Xu R, et al. (2024) Blockade of HMGB1 reduces inflammation and pruritus in atopic dermatitis by inhibiting skin fibroblasts activation. Int Arch Allergy Immunol 185(2): 170–181.
-
He M, Wang J, Zhang J, Li Z, et al. (2023) Inhibition of macrophages inflammasome activation via autophagic degradation of HMGB1 by EGCG ameliorates HBV- induced liver injury and fibrosis. Front Immunol 14: 1147379.
-
Meng X Y, Zhang X, Wang Y, Wang C, Ma W, et al. (2016) EGCG in green tea induces aggregation of HMGB1 protein through large conformational changes with polarized charge redistribution. Sci Rep 6: 22128.
-
Kuang X, Huang Y, Gu HF, Zu XY, Zou WY, et al. (2012) Effects of intrathecal epigallocatechin gallate, an inhibitor of Toll-like receptor 4, on chronic neuropathic pain in rats. Eur J Pharmacol 676(1-3): 51–56.
-
Yang Y, Wang Y, Zhang Y, Li S, Liu J, et al. (2014) Resveratrol reduces the proinflammatory effects and lipopolysaccharide-induced expression of HMGB1 and TLR4 in RAW264.7 cells. Cell Physiol Biochem 33(5): 1283–1292.
-
Dong W-W, Wang X, Bi G, Liu Y, Ma X, et al. (2015) Lung endothelial barrier protection by resveratrol involves inhibition of HMGB1 release and HMGB1- induced mitochondrial oxidative damage via an Nrf2- dependent mechanism. Free Radic Biol Med 88: 404– 416.
-
Karuppagounder V, Arumugam S, Thandavarayan RA, Sreedhar R, Harima M, et al. (2015) Modulation of HMGB1 translocation and RAGE/NF-κB cascade by quercetin treatment mitigates atopic dermatitis in NC/ Nga transgenic mice. Exp Dermatol 24(6): 418–423.
-
Tombulturk FK, Cakmak G, Karadeniz A, Demirel R, Unal B, et al. (2022) Topical application of metformin accelerates cutaneous wound healing in streptozotocin- induced diabetic rats. Mol Biol Rep 49(6): 4851–4860.
-
Menon R, Boldogh I, Hawkins HK, Vinta SR, Basra R, et al. (2016) Placental membrane aging and HMGB1 signaling associated with human parturition. Aging (Albany NY) 8(2): 216–230.
-
Yu Y, Tang D, Kang R (2015) Oxidative stress- mediated HMGB1 biology. Front Physiol 6: 93.
- Epithelioid Granuloma; 3cases with Different Clinical Features
- Advancing Representation in Dermatology Clinical Trials: Ethical, Scientific, and Regulatory Imperatives for Inclusion Across all Fitzpatrick Skin Types
- A Case of Atopic Dermatitis with Concurrent Psoriasis Vulgaris: Successful Treatment with Upadacitinib
- Innovation Lifting Eyeshadow: A Synthesis of Makeup and Optical Illusion
- Distinguishing Superficial Actinic Porokeratosis from Actinic Keratosis with UVF Dermoscopy: A Case Report
- Knowledge, Attitude, and Practices towards Leprosy among Nurses around Semarang, Indonesia