Restrain-stress-induced Biochemical and Transmission Electron Microscopic Changes in Albino Rat’s Brain
Restrained-stress in adult male albino rats for 24 hours in continuity resulted in increased lipid peroxidation (maximum 37.03% in cerebellum) and 8-fold increase (statistically significant p<0.001in brainstem) of fluorescent pigment intensity. Greater affinity of darkly stained luxol-fast positive Purkinje neurons suggests decreased metabolic activity as a result of stress. TEM revealed an increase in the incidence of pleomorphic electrodense bodies in neurons of different brain parts. In cerebellum, dense bodies even exhibited fingerprint pattern due to engulfment of damaged cytomembranes subsequent to increased lipid peroxidation. The electronluscent vacuoles located either at or near the periphery or inside the electrondense bodies, were likely to represent the digested material consequent upon the action of action of hydrolytic enzymes. The increase in the number of coated and smooth vesicles and abundance of microtubules in the cytoplasm of neurons of stressed rats exhibited ultrastructural features in support of their being involved in the transfer of membranes and membrane-associated molecules in the intracellular compartments. Increase in GERL-like profiles could also be detected in cingulate cortex, hypothalamus, and cerebellum after 24 hours restrain of rats. Interestingly, GERL displayed continuity with different cellular organelles in cerebellum of stressed rats. These findings supported the concept advocated by Paavola that GERL’s lytic enzymes engulf membranes and trapped lysosomes thus producing aggregate type inclusions of electrodense bodies.
Introduction
The term ‘Stress’ literally refers to the resisting force set up in the body as a result of an externally applied force called a stressor. In its true medical sense stress is essentially the rate of wear and tear in the body, causing certain alterations in the structure and chemical composition of different organs of the body. The rate of wear and tear, according to Hasan and Glees [1], determines the rate of ageing of an organism. Thus, stress and ageing process are closely akin, if not identical. Under stress, an animal adapts internally and /or externally to meet the requirements due to a particular stressor. Stressors according to Yates [2] may be divided into: (i) physiological stressors – such as hunger, thirst, infection, exhaustion, cold, and heat etc.; and (ii) laboratory stressors – such as shock, shaking, and immobilization or restraint which threaten but are harmless.
Restraint, one of the most commonly used laboratory stressor. Because of the specific positioning of the animal restraint or immobilization causes isometric activity, and muscle cramps. In addition, this stressor is also compounded of fear and is often complicated by voluntary or involuntary fasting, despite a wealth of descriptive literature, using restraint is available in thymus and stomach [3], stomach [4, 5, 6, 7], and testis [8] – there is no report on the effect of restraint stress of the ultrastructural alterations in the brain. Hence the present study was planned to notice the biochemical and transmission electron microscopic effects of restraint in the main parts of the rat brain.
Material and Methods
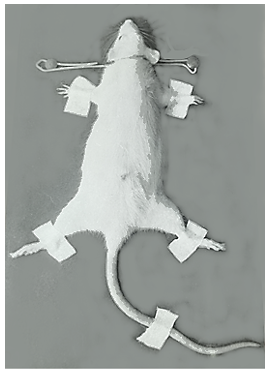
In the present study, 30 healthy male albino rats weighing between 150 to 160 g, from the Central Drug Research Institute, Lucknow. The female rats were purposely excluded from the present experimental study because of their varying cyclic hormonal variations. The rats were housed singly in plastic cages and were fed on standard pellet diet with tap-water ad libitum. The rats were divided equally into control and experimental groups. The rats of the experimental group were immobilized individually as per method advocated by Gottesfeld, et al. [9] by keeping the rat in a prone position on a wooden board by inserting their head though thick aluminium wire-loop (Figure 1) and fastening their all four limbs and tail to metal strips with adhesive tape. The rats were maintained immobilized for a period of 24 hours, during which, like their ‘controls’, they were allowed to ingest tap-water only.

After the stipulated time, the rats of both groups (control and experimental) the rats were sacrificed by: cervical dislocation (for biochemical study), and perfusion technique (Figure 2) for EM purpose. In the former instance, the brain was quickly exposed and divided sharply into: cerebrum, hypothalamus, cerebellum, and brainstem. For EM study, the small pieces were dissected from different brain parts (but here only cerebellum is reported). Specific methods for estimating lipid peroxidation and fluorescent pigment intensity were followed. For TEM study the piece of cerebellum was further trimmed and processed for embedding in a mixture of epon and araldite. Thereafter the pieces were post-fixed in 2 per cent osmium tetroxide. One- micron survey sections were stained with toluidine; ultrathin sections were obtained with the help of an ultratome, and picked up on uncoated copper grids and stained with uranyl acetate [10] and lead citrate [11].
Results
Lipid Peroxidation
The rate of lipid, measured as nano moles of malonaldehyde (MDA) formed per gram brain parts per 120 minutes, in control and in restrained rats is shown in Table 1 and Figure 3.
- Brain Parts
- Range
- Mean
- S.D. ± p. value
- Cerebrum
- Control
- 78.81 – 105.07
- 92.92
- 11.14
- < 0.001
- Stressed
- 107.01 – 129.20
- 118.95
- 7. 68
- Hypothalamus
- Control
- 61.49 – 84.32
- 74.58
- 9.94
- < 0.001
- Stressed
- 89.38 – 105.74
- 100.15
- 7.75
- Cerebellum
- Control
- 71.21 – 95.80
- 84.38
- 10.15
- < 0.001
- Stressed
- 106.92 – 123.81
- 115.63
- 7.3
- Brainstem
- Control
- 46.61 – 78.11
- 62.6
- 10.72
- < 0.001
- Stressed
- 82.61 – 119.25
- 93.62
- 13.78
Table 2: Rate of Lipid peroxidation in different parts of
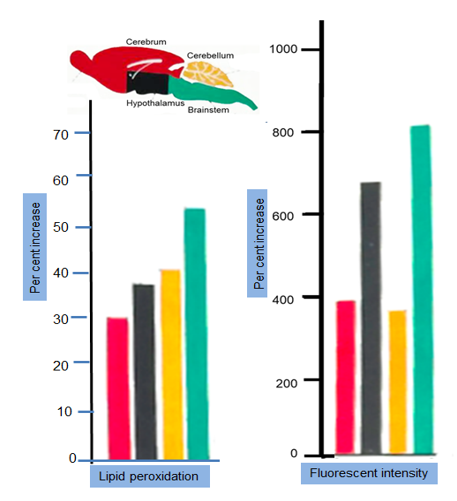
Result: Significant increase in the rate of lipid peroxidation (Figure 4) was observed in the brainstem (49.55%) followed by cerebellum (37.03%), hypothalamus (34.29%), and cerebrum (28.01%).
- Fluorescent Pigment Intensity
- The fluorescent pigment intensity in different parts of control and restrained rats is shown in Table 2, and depicted in Figure 3.
- Brain Parts
- Range
- Mean
- S.D. ± p. value
- Cerebrum
- Control
- 1.0 – 1.7
- 1.28
- 0.28
- < 0.001
- Stressed
- 5.8 – 7.9
- 6.35
- 0.42
- Hypothalamus
- Control
- 0.6 – 1.0
- 0.8
- 0.15
- < 0.001
- Stressed
- 6.0 – 6.7
- 6.35
- 0.3
- Cerebellum
- Control
- 1.0 – 1.8
- 1.4
- 0.3
- < 0.001
- Stressed
- 6.0 – 6.9
- 6.5
- 0.38
- Brainstem
- Control
- 0.9 – 2.1
- 1.6
- 0.19
- < 0.001
- Stressed
- 13.8 – 15.5
- 14.95
- 0.6
Table 1: Fluorescent pigment in different parts of control
Result: A highly significant increase could be observed after 24 hours of restraint stress in all parts of brain studied. There was a 9- fold increase in brainstem; 5-fold increase in cerebrum. There was a regional heterogeneity of the per cent increase in the fluorescent intensity in various brain parts studied.
Microscopic Findings
Only those observed in cerebellum are reported in the present communication
Survey Sections (One-micron Toluidine Blue- stained)
Sections of cerebellum stained with luxol fast blue and toluidine methods, revealed under higher magnification, the presence of two types of Purkinje neurons: the light and dark stained (Figure 4) on the basis of their differential stainability. The darker or luxol fast positive cells had clearly defined cell boundaries and were somewhat smaller than lighter luxol fast negative cells. Interestingly, the luxol positive Purkinje cells increased in distribution as compared to those in control rats.
![Figure 4: Survey section of cerebellar hemisphere cortex in- [a] control and [b] immobilized rat shows relatively more numbers of dark (metabolically inactive Luxol positive) Purkinje neurons in stressed rats after 24hours of restraint.](/fulltextimages/13091/fig_4.jpeg)
(a) Control rat (Luxol blue stain x 920)-more light [3] purkinje cells and 1 dark cell.
(b) Immobilized rat (Luxol blue stain x 920)- more dark [3] purkinje cells and 1 light cell.
Ultrathin Transmission Electron Microscopic Sections
In ‘Control’ rats: The molecular layer of cerebellar cortex revealed abundant nerve fibres, majority of which were unmyelinated with only a few sparse at places. Purkinje neurons were characterised by nuclear membrane infolding. They had light finely granular euchromatin. A few patches of darker-stained heterochromatin were also noticeable. Nucleoli were infrequently visible. Mitochondria exhibited pleomorphic shapes mostly with elongated profiles and well-defined tubular cristae parallel to one another. Long profiles of rough endoplasmic reticulum were prominently seen.
In ‘Stressed’ rats: The molecular layer of stressed rat did not show any appreciable change as compared with control rats. However, the Oligodendrocytes, at places were characterized by the presence of many electrondense polymorphic granules in their cytoplasm.
The Purkinje neurons consisted of infoldings of the nuclear membrane. Pleomorphic mitochondria were seen with highly electrondense matrix. A number of mitochondria lied very close to electrondense bodies (Figure 5). At higher magnification, some mitochondria had disorganized cristae. RER consisted of tubular cisternae seen mainly in close proximity of the nuclear membrane, and were studded on both sides
![Figure 5: Transmission electron micrograph of cerebellum of a stressed rat. EDB – electron dense body [lipofuscin], GERL- Golgi endoplasmic reticulum lysosome complex; M- mitochondria; RER- rough endoplasmic reticulum with ribosomes.](/fulltextimages/13091/fig_5.png)
However, short profiles of tubular cisternae invested with electrondense material as noted in control animals were not observed in any stressed rat. Ribosomes seen mainly in the vicinity of the nuclear membrane were relatively decreased. Golgi complexes had characteristic prominent dilatations at the ends of their cisternae arranged in stacks. The coated and uncoated vesicles were remarkably increased. Majority of coated vesicles exhibited a typical ‘cart-wheel’ appearance. Long profiles of microtubules were observed in abundance. There was an appreciable increase in number and size of membrane-bound eletrondense bodies. Such bodies were pleomorphic in shape. A few electrondense bodies were noticed in the vicinity of degenerating mitochondrial profiles (Figure 5). Numerous GERL-like profiles could be observed throughout the cytoplasm of stressed rat cerebellum. The granule cell layer appeared changeless.
Discussion
Lang and Vessel [12] emphasized that a number of environmental factors; such as handling methods of animals, cage-type, bedding material, noise pollution, light-and dark periods (photoperiods) and some other parameters significantly modify animals’ behaviour, physiological, and biochemical responses. Keeping the above factors into consideration, the rats selected for the present study were procured every time from the same colony maintained in the animal house of the Lucknow situated Central Drug Research Institute (CDRI). 24 hours prior to start the experiment, the control as well stress group of albino rats were deprived of any food except tap-water. Immobilization model of laboratory stressor was selected as per method advocated by Gottesfeld, et al. [9]. Confirmation of gastric mucosa ulceration in stressed rats after 24 hours was indicator for successful stress-model.
An enhanced lipid peroxidation in all parts studied during 24 hour restrain-stress in rats in vivo has been earlier identified [13] as a basic deteriorative reaction in cellular mechanisms. Stimulated fluorescent pigment intensity observed in the present investigation was correlated directly with the extent of increased lipid peroxidation. Characterization of Lipofuscin pigment reveals that they are complexes of lipid protein substances of polyunsaturated lipids of subcellular membranes.
The increase in the density of dark Purkinje (luxol blue positive) neurons in stressed rats was suggestive of a remarkable change in the cerebellar histology. A similar finding of Meitner [14] has been reported by the author after 96 hours immobilization of rats. Furthermore, Meitner [14] also reported a significantly more dendritic branching in Purkinje neurons after stress. This finding could not be substantiated in this study.
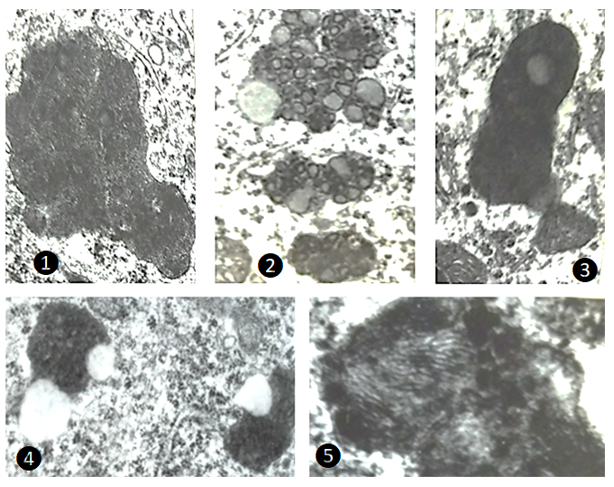
The most consistent alteration detected in various regions of rat brain after 24-hour immobilization stress was the increased incidence of pleomorphic electrondense bodies in the neurons. Strikingly, regional heterogeneity in the fine structure of these bodies was noticed (Figure 6). It was proposed long ago by Dolley [15] that chromatin substance extruded from the nucleus into the neuronal cytoplasm was possibly transformed into pigment granules. Hasan and Glees [16] propounded a hypothesis that, “… a chain of activity, starting in the vicinity of the nucleus perhaps linked with DNA-RNA turnover and associating Golgi apparatus, endoplasmic reticulum, lysosomes and mitochondria causes the ultimate deposition of Lipofuscin pigment in the cytoplasm”.
![Figure 6: Transmission electron microscopic appearances of pleomorphic electrodense bodies in cerebellum [1 and 2; x22,400] and certain other brain parts such as amygdaloid complex [4; x15000] and cingulate cortex of cerebrum [3; x25000 and 5; x 49000].](/fulltextimages/13091/fig_6.png)
The present study is possibly the first account to describe the presence of structures which correspond to GERL in the neurons of different parts of the brain of adult male rats immobilized for 24 hours in continuity. It is universally accepted view that GERL is a hydrolase-rich region of endoplasmic reticulum, spatially related to Golgi apparatus (but not a part of it). It is stipulated that even though the pathway to GERL is complicated it may be summarized that GERL appeared to participate in the formation of lytic bodies and thus ultimately to form electrondense bodies (Figure 7) which are most likely as Lipofuscin pigment (commonly termed as wear-and-tear pigment).
On the basis of the criteria for identifying GERL profiles, the present study revealed a significant increase in GERL profiles, and coated and smooth vesicles derived from GERL. Even though kinetic data are lacking, the evidences from different parts of the rat brain in the present study suggest that the stress-induced electrondense bodies (? lipofuscin) are mainly derived from GERL. Also, the close proximity of many coated vesicles with GERL is indicative that the former was budded off from smooth-surfaced cisternal elements of GERL. The present concept of lipofuscin genesis has the support of an earlier report by Paavola [17] and more recent evidence put forward by Bouldin, et al. [18].
Conclusions
Restrained-stress in adult male albino rats for a period of 24 hours in continuity was noticed to cause the following biochemical and ultrastructural alterations in different parts of the brain:
- Increased lipid peroxidation was due to oxidation of unsaturated fatty acids in brain parts: 49.55% maximum in brainstem followed by cerebellum (37.03%), hypothalamus (34.29%), and cerebrum (28.01%).
- Brainstem appeared to be most vulnerable target after restraint. A statistically significant increase of nearly 8-fold intensity of fluorescent pigment.
- Some cerebellar Purkinje neurons depicted a significantly greater staining affinity for luxol fast blue stain. The darkly stained luxol-positive cells were suggestive of their decreased metabolic activity due to immobilization stress.
- Despite of statistically significant (p<0.
5. TEM findings revealed an increase in the incidence of pleomorphic electrodense bodies in neurons of different brain parts. In cerebellum, restraint-induced dense bodies even exhibited fingerprint pattern at higher magnifications. These dense bodies were suggestive of the lysosomal involvement in engulfing damaged cytomembranes subsequent to lipid peroxidation as a result of immobilization stress 6. The electronluscent vacuoles located either at or near the periphery, or inside the electrondense bodies, were interpreted to represent the digested material consequent upon the action of action of hydrolytic enzymes. The lucent vacuoles were considered likely to be composed of saturated lipids which could not react with osmium tetroxide during the processing of tissues to prevent the electrondense appearance. 7. The increase in the number of coated and smooth vesicles and abundance of microtubules in the cytoplasm of neurons of stressed rats exhibited ultrastructural features in support of their being involved in the transfer of membranes and membrane-associated molecules in the intracellular compartments. 8. Increase in GERL-like profiles could also be detected in cingulate cortex, hypothalamus, and cerebellum after 24 hours restrain of rats. Interestingly, GERL displayed continuity with different cellular organelles in cerebellum of stressed rats. These findings supported the concept advocated by Paavola (1978) that GERL’s lytic enzymes engulf membranes and trapped lysosomes thus producing aggregate type inclusions of electrodense bodies.
GERL profiles observed in the present investigation may be as a result of stress-induced secretory proteins that are reported in the literature as: (a) synthesized on ribosomes associated with the endoplasmic reticulum; (b) segregated within the cisternal space of the rough endoplasmic reticulum; (c) transported to the vicinity of the Golgi apparatus; (d) concentrated and further processed in Golgi saccules or condensing vacuoles; (e) stored in secretory granules; and (f) discharged from secretory granules to the outside of the cell. Although much is known about the secretory process, many specific details noticed after 24-hour restraint stress remain speculative. Included among the poorly understood aspects of the secretory process are the roles of the Golgi apparatus and/or GERL’ in the processing and packaging of secretory proteins after restraint stress.
References
-
Hasan M, Glees P (1972a) Genesis and possible dissolution of neuronal lipofuscin. Gerantologia 18: 217- 236.
-
Yates EF (1967) The Adrenal Cortex. Little Brown and Co., Boston, USA, pp: 133-184
-
Selye H (1936) Thymus and adrenals in response of the organism to injuries and intoxications. Brit J Exp Pathol 17: 234-248.
-
Rossi G, Bonfils S, Lieffoogh F, Lambling A (1956) Technique nouvelle pour produire des ulcerations gastriques chez le rat blanc. Compt Rend Soc Biol 150: 2124-2126.
-
Brodie DA, Hanson HM (1960) A Study of the Factors Involved in the Production of Gastric Ulcers by the Restraint Technique. Gastroenterology 38(3): 353-360.
-
Parisio C, Clementi F (1976) Surface alterations induced by stress in gastric mucosa: Protective effect of Zolimidine. A transmission and scanning electron microscope investigation. Lab Invest 35(5): 484-495.
-
Kholkute SD, Udupa (1978) Effect of immbobilization stress on implantation and pregnancy in rat. Indian J Exp Biol 16: 799-800.
-
Kholkute SD, Udupa KN (1979) Effect of immbobilization stress on spermatogenesis and accessory sex organs in rats. Indian J Exp Biol 17: 206-208.
-
Gottesfeld Z, Kvetnansky R, Kopin IJ, Jacobowitz M (1978) Effects of repeated immbobilization stress on glutamate decarboxylase and choline acetyltransferase in discrete brain regions. Brain Res 152: 374-378.
-
Watson ML (1958) Staining of tissue sections for electron microscopy with heavy metal. J Biophys Biochem Cytol 4: 475-478.
-
Reynolds ES (1963) The use of lead citrate at high pH as an electron opaque stain in electron microscopy. J Cell Biol 17: 208-212.
-
Lang CM, Vessel ES (1976) Experimental and genetic factors affecting laboratory animals: Impact on biomedical research. Fed Proc 35(5): 1123-1124.
-
Tappel AL (1973) Lipid peroxidation damage to cell components. Fed Proc 32(8): 1870-1874.
-
Meitner ER (1977) Difference in the colorability of Purkinje cells. Acta Anat 97(2): 191-199.
-
Dolley DH (1917) The recovery from depression in the Purkinje cell and the decline to senility of depression with the histogenesis of abnormal pigmentation. J Comp Neurol 28(3): 465-493.
-
Hasan M, Glees P (1972b) Electron microscopical appearance of neuronal Lipofuscin using different preparative techniques including freeze-etching. Exp Geront 7(5): 345-351.
-
Paavola LG (1978) The corpus luteum of the guinea pig. III. Cytochemical Studies on the Golgi Complex and GERL during Normal Postpartum Regression of Luteal cells Emphasizing the Origin of Lysosomes and Autophagic Vacuoles. J Cell Biol 79(1): 59-73.
-
Bouldin TW, Goines ND, Bangell CR, Krigmann MR (1981) Pathogenesis of trimethltin neuronal toxicity. Am J Pathol 104(3): 237-249.
- Pattern of Breast Lesions in Ovu Inland, Delta State, South Southern Nigeria
- Morphometric Analysis of the Human Femur: Exploring Platymetric and Robusticity Indices Among the Nigerian Population
- Anatomical Variation of Arteria Lusoria: Clinical Implications for Dysphagia Lusoria and Surgical Risk
- Morphometric Study of the Vertebral Body and Pedicle of Typical Cervical Vertebrae Using Radiological Image
- Epigenetic Mechanisms Driving Human Evolutionary Changes
- Neuroprotective Effects of Ginkgo Biloba Extract on Bilateral Common Carotid Artery Ischaemic Stroke Induced in Wistar Rat