The Applicability of Genetic Markers Based on Molecules Micro RNA in Agricultural Research
Within our research we are focused on the applicability and exploitation of genomic markers based on microRNA (miRNAs) molecules for purposes of genotyping and genomic activity mapping of miRNA-based markers. The miRNAbased genotyping systems have been applied in the germplasm evaluation of flax (Linum usitatissimum L.). Obtained results have shown that miRNA-based molecular markers are effective for the evaluation of flax genome polymorphism in the context of the origin as well as for mapping the activity of miRNAs molecules during different developmental stages of various plant tissues.
Ražná K* and Hlavačková L
Slovakia
mail: katarina.razna@uniag.sk various plant tissues.
Keywords: Genetic Markers; MicroRNA; Genotype; Linum usitatissimum L; QRT-PCR analysis
Introduction
Endogenous, non-coding and hairpin-derived molecules microRNAs (miRNAs) are about 20–24 nucleotides long. They control gene expression by binding to the target mRNAs, leading to either translation delay or mRNA degradation [1]. Transcription factors represent the main target sequences of microRNA molecules [2, 3]. MiRNAs are involved in wide variety of biological and metabolic pathways of plants. They represent a vital part in developmental and physiological processes and affect the plant genome response to environmental stress. Many stress-regulated genes are found to be regulated by miRNAs. MicroDNA sequences are common in the plants genomes however most annotated miRNAs are located in intergenic regions [4, 5]. The high conservation of miRNA sequences provided an opportunity to develop an effective type of molecular markers that is useful for studying of genetic diversity [6]. The miRNA-based molecular markers have been reported in very few plants; Brassica species [4], Setaria italica L. [7] and Oryza sativa L. [8, 6]. The advantages of microRNA-based marker system including high polymorphism and reproducibility, transferability across species and putative functionality [4, 9, 10]. The polymorphism level is comparable with SSR markers [4]. Despite large number of available SSRs markers, most of them originated from protein-coding or untranslated regions [6]. Sequences microRNAs represent conserved, non-coding sequences with the regulatory function of many biological processes. Most of the fragments are produced between 30 bp and 330 bp (occasionally up to 500 bp) in length. Individual fragment sizes can be clearly distinguished by agarose and polyacrylamide gel electrophoresis. Marker assay primers are design based on the sequences of mature miRNAs which are part of the step-loop structure. Therefore, the specificity of these markers relies on the capability of miRNA-based primers to amplify different parts of specific stem-loop structure as well as variable length polymorphism of regions between neighboring miRNAs molecules. In addition, miRNA-based primers can be combined randomly (it means combinations of various forward primers or forward and reverse primers) which it is not possible in the case of SSR primers [4, 9]. However, miRNA-based microsatellite markers have been developed from salt responsive miRNA genes for genetic diversity analysis of salt responsive miRNAs among the salt tolerant and sensitive rice genotypes of rice [6, 8]. Another approach includes the integration of amplified fragment length polymorphism (AFLP) primers with primers designed from conserved miRNA genes. This marker system comprising of qRT-PCR analysis, is focused on the polymorphism of miRNA genes which provides a reliable targeted genotyping strategy to assess genetic diversity with regard to miRNA gene variation and evolution. The potential of miRNA markers as functional markers has been confirmed by the study of Htwe et al. (2015) which reported miR1511-InDel as a soybean- specific functional marker for the study of genetic diversity, genotyping of germplasm and evolution studies. The presence of allelic variation within the precursor pre- miR1511 in soybean germplasm has allowed developing miR1511-InDel marker that could be distinguished on the presence or absence of miR1511.
Material and Methods
The seed material has been provided by the N. I. Vavilov Research Institute of Plant Genetic Resources St. Petersburg (Russia) and by company AGRITEC, Research, Breeding and Services, Ltd. from Šumperk (Czech Republic). Genotyping analyses were carried out on eight selected flax genotypes of various origin and agronomical type (Table 1) by three combinations of miRNA-based primers. Seven out of eight genotypes are of commercial types of L. usitatissimum L. and one genotype of L. crepitans as a wild type of flax. The selected genotypes were classified into subspecies based on the classification by E.V. Elladi. For the purpose of mapping activities of miRNAs molecules during developmental stages connected to seed formation were selected eight genotypes mostly of intermediate agronomical type (Table 2). In order to consider the level of alpha-linolenic acid content in the seeds, two genotypes of linseed type have been included. These genotypes have been analyzed by two combinations of miRNA-based primers within three different developmental stages and three types of plant tissues. Samples were taken from buds, flowers petals and bolls in the following phenological stages: buds - BBCH 55 (first individual flowers visible- still closed), flowers petals - BBCH 65 (full flowering: 50% of flowers opened), bolls- BBCH 71 (10% of bolls have reached final size).
| Genotype | |||||||||||||||||
| Code | Species | Subsp. | Type | Origin | |||||||||||||
| designator | |||||||||||||||||
| 1 | kk-1127 | L. usitatissimum | mediterraneum | Intermediate | Tunisia | ||||||||||||
| 2 | kk-1195 | L. usitatissimum | indo-abyssinicum | Linseed | Ethiopia | ||||||||||||
| 3 | k-1288 | L. usitatissimum | eurasiaticum | Linseed | Afghanistan | ||||||||||||
| 4 | k-3002 | L. usitatissimum | hindustanicum | Intermediate | India | ||||||||||||
| 5 | k-3730 | L. usitatissimum | eurasiaticum | Intermediate | China | ||||||||||||
| 6 | k-7015 | L. usitatissimum | mediterraneum | Linseed | Tunisia | ||||||||||||
| 7 | k-7130 | L. usitatissimum | mediterraneum | Intermediate | Morocco | ||||||||||||
| 8 | u-562348 | Linum crepitans | - | Linseed | Portugal |
Table 1: List of flax genotypes used in the genotyping analyzes by miRNA-marker assay.
| Code | Name | Type | Origin | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Amon | Intermediate | Czech Republic | ||||||||
| 2 | Libra | Linseed | Czech Republic | ||||||||
| 3 | Raciol | Linseed | Czech Republic | ||||||||
| 4 | Lin 225 | Intermediate | Netherlands | ||||||||
| 5 | Bolley Golden | Intermediate | USA | ||||||||
| 6 | Currong | Intermediate | Australia | ||||||||
| 7 | Mermilloid | Intermediate | Czech Republic | ||||||||
| 8 | Eyre | Intermediate | Australia |
Table 2: List of flax genotypes used in the analyzes for mapping activities of miRNA-based markers.
The total genomic DNA extraction was performed based on Padmalantha and Prasad (2006) [11]. After extraction DNA followed by quantification (Implen NanoPhotometer®), the purity and integrity assessment by the absorbance at 260/280 nm ratio, was DNA diluted to 70 ng.μl-1. The primers for the miRNA-based markers were designed according to the mature miRNAs sequences of flax, originating from the miRNA database (http://www.mirbase.org/) considering the principles of primer design [4, 7]. The single forward primers and the universal miRNA reverse primer [12, 13] were combined to perform a marker assays (Table 3). Due to the species transferability of miRNA-based primers was possible to confirmed single forward primers with each other. The effectiveness and species transferability of used primers was in previous studies [9, 10, 14, 15, 16].
| Primer name | Sequences |
|---|---|
| miR156b-F | 5´-TGACAGAAGAGAGAGAGCACA-3´ |
| miR-R | 5´-CCAGTGCAGGGTCCGAGGTA-3´ |
| mir168a-F | 5´-CACGCATCGCTTGGTGCAGGT-3´ |
| miR-R | 5´-CCAGTGCAGGGTCCGAGGTA-3´ |
| miR408a-F | 5´-GGCTGGGAACAGACAGAGCATGGA- 3´ |
| miR-R | 5´-CCAGTGCAGGGTCCGAGGTA-3´ |
| miR156b-F | 5´-TGACAGAAGAGAGAGAGCACA-3´ |
| miR408a-F | 5´-GGCTGGGAACAGACAGAGCATGGA- 3´ |
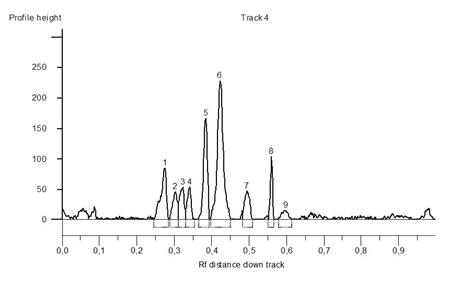
miRNA-based markers were PCR amplified in a 20-μl reaction mixture that contained 70 ng of genomic DNA, 1 × DreamTaq Buffer (KCl, (NH4)2SO4, 20 mmol.dm-3 MgCl2), 2 units of DreamTaq DNA polymerase, 0.8 mmol.dm-3 dNTPs (Bioline), 10 pmol.dm-3 of each primer and nuclease-free water for PCR amplification. The PCR amplification program used the ‘touchdown’ method as follows: initial denaturation at 94°C for 5 min; 5 cycles of 30 s at 94°C, 45 s at 64°C (annealing temperature was decreased with 1°C/cycle), and 60 s at 72°C; 30 cycles of 30 s at 94°C, 45 s at 60°C, and 60 s at 72°C; and a final extension at 72°C for 10 min. PCR products were separated on 15% TBE-Urea gels (Invitrogen) running in 1 × TBE Running Buffer at a constant power 180 V, 30 mA for 75 min. The polyacrylamide gels were stained with GelRedTM Nucleic Acid Gel stain and were visualized on G-Box Syngene electrophoresis documentation system. For the recording of loci number and unique identification of fragments, the gels were analyzed by the GeneTools software (Syngene) (Figure 1). Each fragment is characterized by quantity and volume profile in pixels. Profiles are recorded on the basis of the set threshold value in which the analysis is carried out. Thus, it is possible to record the fragments which are not visually clearly observable.
Results and discussion
We have investigated the activity of specific miRNAs families of flax (Linum usitatissimum L.) in the context of the genotype origin and depending on plant tissues and developmental stages. The research has been conducted in order to apply selected types of miRNA-based markers of flax, miR156b, miR168a, miR408a and their combination (miR156b/miR408a) for the genotyping and miRNA-based markers activity mapping analysis. Selection of primers was done according to the purpose of the research and target sequences of microRNA molecules. The miRNA network which includes the miR156 family belongs to one of the extensively reviewed. The miR156 targets the squamosa promoter binding protein (SBP), transcription factor, which is involved in controlling the flowering time and transition from the juvenile to adult vegetative phase in monocot and dicots [2]. The miR408a of Linum usitatissimum L., is included in the regulation of metabolism and transport processes and its expression is tissue specific [2, 3]. The high level of expression indicates that it might play an important role in flax development [2]. However, miR408a is considered as potential biomarker in plant stress response [17, 18]. One of the targets of miR168 family is sequences of cytochrome P450 which is involved in a wide range of biosynthetic reactions, including fatty acid biosynthesis. The miR168 is also considered as the biomarker of plant stress response [17]. The highest expression of flax miR168 is exhibited in anthers and flowers [2]. A total of 145 miRNA loci were identified, of which 19 were unique. By the miR156b-F/miR-R primers pair was amplified 39 miRNA loci, out of which 6 were unique (Figure 2A). Each of eight accessions had their own unique profile of the miRNA loci, which allowed differentiating these genotypes based on their miRNA fragments fingerprinting pattern. By miR408a-F/miR-R primer pair, were amplified 49 miRNA loci in total, out of which 4 were unique (Figure 2B). The unique miRNA loci were recorded in the oily genotype from Ethiopia and two intermediate genotypes, one from India and one from China, while these two genotypes were also characterized by a significantly different miRNA fingerprinting profile. The polymorphism profile of L. crepitans as a representative of wild type of flax included the unique miRNA loci determined by the combination of miR156b- F/miR-R and miR156b-F/miR408a-F primer. By the combination of two applied primers, miR156b and miR408a (picture not shown) were amplified 57 miRNA loci, out of which 9 were unique. Each of the genotypes was characterized by a specific miRNA profile. From the applied miRNA-based molecular markers, the combination of miR156b-F/miR-R and miR156b- F/miR408a-F primer was proven to be more suitable for this type of analysis.
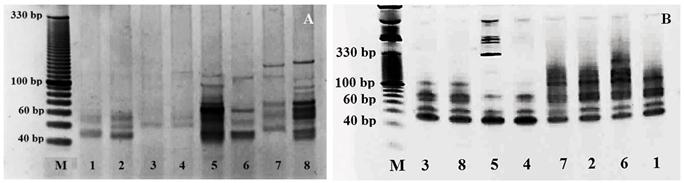
Figure 2: PCR amplification profiles generated with miR156b-F/miR-R (A) and miR408a-F/miR-R (B) primers pair in flax genotypes from 1 - Tunisia (intermediate type), 2 - Ethiopia, 3 - Afghanistan, 4 - India, 5 - China, 6 - Tunisia (linseed type), 7 - Morocco, 8 - Portugal. Because most of the microRNA molecules play important role in the regulation of developmental processes, we were interested in the profile activity of miRNA not only during different developmental stages but also in various plant tissues. For these purposes were selected two categories of conserved miRNA families miR156 and miR168. The genomic analyzes have been carried out under different development stages and various organs and tissues (flower buds, flowers, bolls, leaves, seeds). Samples for analyzing polymorphism were taken from buds, flowers petals and bolls in the following phenological stages: buds, flowers petals and bolls.
The representation of miR168 loci in the flax genome have increased according to developmental stages ascending (stage of bud, flower and boll). Cytochrome P450, one of the target sequences of miR168 is involved in the formation of fatty acid precursors. Taken into account the abundancy of miRNAs which is correlated with the level of expression of their target sequences [2, 3] is possible to assume that some of the biosynthetic processes take place during the formation of flower bud. The number of miRNA loci amplified by miR156-based molecular marker as well as the extent of PCR amplification reached lower values in comparison to miRNA168 based molecular marker. However, almost in all genotypes it was possible to observe the increase of miRNA156b loci amplification (Figure 3).
![Figure 3: Number of amplified miR156 loci of eight genotypes depending on developmental stage of flax. The lower representation of miR156 loci, in the phases of flower buds formation and flowering, complies with the results of Barvkar et al. (2013) [2] concerning an inverse relation between the expression of the miRNAs and their putative target gene, squamosa promoter-binding protein (SBP protein). Low miRNA profile in these stages is pointing toward higher expression of targeted sequences in flowers and flower organs which role is to control flowering time and the transition from the juvenile to the adult vegetative phase. Taking into account the genetic background of genotypes, was possible to identified unique loci of linseed and intermediate genotypes within individual developmental stages (Figure 4). Different situation could be observed in the case of linseed genotype with code 3 (Raciol, characterized by high content of alpha-linolenic acid). The miRNA loci amplification depending on the development stage gradually decreased, indicating the shift of fatty acids metabolism towards seed development stages. The abundance of mature miRNAs, which is in direct correlation to the expression of MIRNA genes, varies considerably depending on the type of miRNAs family, types of tissue or developmental stages, suggesting the spatially and temporally regulated expression patterns of plant miRNAs [19].](/fulltextimages/459/fig_3.png)
Figure 3: Number of amplified miR156 loci of eight genotypes depending on developmental stage of flax. The lower representation of miR156 loci, in the phases of flower buds formation and flowering, complies with the results of Barvkar et al. (2013) [2] concerning an inverse relation between the expression of the miRNAs and their putative target gene, squamosa promoter-binding protein (SBP protein). Low miRNA profile in these stages is pointing toward higher expression of targeted sequences in flowers and flower organs which role is to control flowering time and the transition from the juvenile to the adult vegetative phase. Taking into account the genetic background of genotypes, was possible to identified unique loci of linseed and intermediate genotypes within individual developmental stages (Figure 4). Different situation could be observed in the case of linseed genotype with code 3 (Raciol, characterized by high content of alpha-linolenic acid). The miRNA loci amplification depending on the development stage gradually decreased, indicating the shift of fatty acids metabolism towards seed development stages. The abundance of mature miRNAs, which is in direct correlation to the expression of MIRNA genes, varies considerably depending on the type of miRNAs family, types of tissue or developmental stages, suggesting the spatially and temporally regulated expression patterns of plant miRNAs [19].
![Figure 4: PCR amplification profiles generated by primers pair miR156b-F/miR-R and miR168a-F/miR-R across buds tissues (A and B), flowers petals tissues (C and D) and bolls tissues (E and F). Legend: M - 10 bp DNA Ladder Invitrogen, 1- Amon, 2- Libra, 3- Raciol, 4- Lin 225, 5- Bolley Golden, 6- Currong, 7- Mermilloid, 8- Eyre. DNA-based molecular markers are an integral part of the evaluation and assessment of Genomic significance of plant genetic resources. Our results confirmed the stability, polymorphism, functionality and transferability potential of miRNA-based markers, for genotyping analyses, which corresponds to the results of several studies of Fu et al. (2013), Yadav et al. (2014), Mondal and Ganie (2014) and Ganie and Mondal (2015) [4,6-8]. Success of transferability of these type of markers relies on the fact that since these markers are derived from the sequences of mature miRNAs, the high degree of cross- transferability across the genera is expected [7]. Our observations have also supported the role of miRNA molecules as potential biomarkers of abiotic stress [17]. This indicated a highly efficient miRNA-based marker system offers a wide range of applications in agricultural research. DNA-based molecular markers are an integral part of the evaluation and assessment of genomic significance of plant genetic resources. Our results confirmed the stability, polymorphism, functionality and transferability potential of miRNA-based markers, for genotyping analyses, which corresponds to the results of several studies of Fu et al. (2013), Yadav et al. (2014), Mondal and Ganie (2014) and Ganie and Mondal (2015) [4,6-8]. Success of transferability of these types of markers relies on the fact that since these markers are derived from the sequences of mature miRNAs, the high degree of cross-transferability across the genera is expected [7]. Our observations have also supported the role of miRNA molecules as potential biomarkers of abiotic stress [17]. This indicated a highly efficient miRNA-based marker system offers a wide range of applications in agricultural research.](/fulltextimages/459/fig_4.jpeg)
Figure 4: PCR amplification profiles generated by primers pair miR156b-F/miR-R and miR168a-F/miR-R across buds tissues (A and B), flowers petals tissues (C and D) and bolls tissues (E and F). Legend: M - 10 bp DNA Ladder Invitrogen, 1- Amon, 2- Libra, 3- Raciol, 4- Lin 225, 5- Bolley Golden, 6- Currong, 7- Mermilloid, 8- Eyre. DNA-based molecular markers are an integral part of the evaluation and assessment of Genomic significance of plant genetic resources. Our results confirmed the stability, polymorphism, functionality and transferability potential of miRNA-based markers, for genotyping analyses, which corresponds to the results of several studies of Fu et al. (2013), Yadav et al. (2014), Mondal and Ganie (2014) and Ganie and Mondal (2015) [4, 6, 7, 8]. Success of transferability of these type of markers relies on the fact that since these markers are derived from the sequences of mature miRNAs, the high degree of cross- transferability across the genera is expected [7]. Our observations have also supported the role of miRNA molecules as potential biomarkers of abiotic stress [17]. This indicated a highly efficient miRNA-based marker system offers a wide range of applications in agricultural research. DNA-based molecular markers are an integral part of the evaluation and assessment of genomic significance of plant genetic resources. Our results confirmed the stability, polymorphism, functionality and transferability potential of miRNA-based markers, for genotyping analyses, which corresponds to the results of several studies of Fu et al. (2013), Yadav et al. (2014), Mondal and Ganie (2014) and Ganie and Mondal (2015) [4, 6, 7, 8]. Success of transferability of these types of markers relies on the fact that since these markers are derived from the sequences of mature miRNAs, the high degree of cross-transferability across the genera is expected [7]. Our observations have also supported the role of miRNA molecules as potential biomarkers of abiotic stress [17]. This indicated a highly efficient miRNA-based marker system offers a wide range of applications in agricultural research.
Acknowledgements
This research was supported by the project by European Community under project no 26220220180: Building Research Centre ‘AgroBioTech’. The authors thank for providing them seed material for research purposes to the N. I. Vavilov Research Institute of Plant Genetic Resources St. Petersburg (Russia) and by company AGRITEC, Research, Breeding and Services, Ltd. from Šumperk (Czech Republic).
References
-
Erson-Bensan AE (2014) Introduction to MicroRNAs in Biological Systems. In Yousef M, Allmer J (Eds.) MicroRNA Biology and Computational Analysis pp. 1- 14.
-
Barvkar VT, Pardeshi VC, Kale SM, Qiu S, Rollins M, et al. (2013) Genome-wide identification and characterization of microRNA genes and their targets in flax (Linum usitatissimum): Characterization of flax miRNA genes. Planta 237(4): 1149-1161.
-
Neutelings G, Fénart S, Lucau-Danila A, Hawkins S (2012) Identification and characterization of miRNA and their potential targets in flax. Int J Plant Physiol 169 (17): 1754-1766.
-
Fu D, Ma B, Mason AS, Xiao M, Wei L, et al. (2013) MicroRNA-based molecular markers: a novel PCR- based genotyping technique in Brassica species. Plant Breeding 132(4): 375-381.
-
Zhang L, Chia JM, Kumari S, Stein JC, Liu Z, et al. (2009) A genome-wide characterization of microRNA genes in maize. PLoS Genet 5(11): e1000716.
-
Ganie SA, Mondal TK (2015) Genome-wide development of novel miRNA-based microsatellite markers of rice (Oryza sativa) for genotyping applications. Molecular Breeding 35(51): 1-12.
-
Yadav CBY, Muthamilarasan M, Pandey G, Prasad M (2014) Development of novel microRNA- based genetic markers in foxtail millet for genotyping applications in related grass species. Molecular Breeding 34(4): 2219-2224.
-
Melnikova NV, Dmitriev AA, Belenikin MS, Speranskava AS, Krinitsina AA, et al. (2014) Identification and characterization of salt responsive miRNA-SSR markers in rice (Oryza sativa). Gene 535(2): 204-209.
-
Ražná K, Hlavačková L, Bežo M, Žiarovská J, Habán M, et al. (2015a) Application of the RAPD and miRNA markers in the genotyping of Silybum marianum (L.) Gaertn. Acta phytotechnica et zootechnica 18(4): 83- 89.
-
Hlavačková L, Nôžková J, Porokhovinova E, Brutch N, Shelenga T, et al. (2016) Analysis of miRNA polymorphism during the selected developmental processes of flax. Journal of Central European Agriculture 17(3): 707-724.
-
Padmalatha K, Prasad MNV (2005) Optimization of DNA isolation and PCR protocol for RAPD analysis of selected medicinal and aromatic plants of conservation concern from Peninsular India. African Journal Biotechnology 5(3): 230-234.
-
Kulcheski FR, Marcelino-Guimaraes FC, Nepomuceno AL, Abdelnoor RV, Margis R (2010) The use of microRNAs as reference genes for quantitative polymerase chain reaction in soybean. Analytical Biochemistry 406(2): 185-192.
-
Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee DH, et al. (2005) Real-time quantification of microRNAs by stem-loop RT-PCR. Nucleic Acids Research 33(20): e179.
-
Hlavačková L, Ražná K (2015) Polymorphism of specific miRNAs in the context of flax (Linum usitatissimum L.) genome adaptability to abiotic stress. MendelNet 416-420
-
Ražná K, Bežo M, Hlavačková L, Žiarovská J, Miko M, et al. (2016) MicroRNA (miRNA) in food resources and medicinal plant. Potravinárstvo 10(1): 188-194.
-
Ražná K, Nôžková J, Hlavačková L, Brutch N, Porokhovinova E, et al. (2015b) Genotyping of flax genetic resources by miRNA-based molecular markers and morphology. Agriculture 61(4): 129- 138.
-
Bej S, Basak J (2014) MicroRNAs: The Potential Biomarkers in Plant Stress Response. American Journal of Plant Sciences 5(5): 748-759.
-
Sunkar R (2010) MicroRNAs with macro-effects on plant stress responses. In Seminars in Cell & Developmental Biology 21(8): 805-811.
-
Xie Z, Khanna K, Ruan S (2010) Expression of microRNAs and its regulation in plants. Semin Cell Dev Biol 21(8): 790-797.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note