Analysis of Gene Expressions Associated with Response of Phenolic Metabolism of Allelopathic Rice under Barnyardgrass Stress
Allelopathic rice varieties Giza 179 and Sakha 103 were induced by exogenous salicylic acid (SA) to suppress barnyardgrass weed (BYG) and were investigated using suppression subtractive hybridization (SSH) and real-time fluorescence quantitative PCR (qRT-PCR) methods. The results explained that exogenous salicylic acid could induce the allelopathic effect of rice on BYG. The variety Giza 179 showed higher inhibitory effect than Sakha 103 variety on BYG after treated with different concentrations of salicylic acid. Fourteen genes induced by salicylic acid were obtained by SSH analysis from Giza 179 variety. These genes encode ubiquitin carrier proteins, proteins related to phenyl propanoid metabolism, receptor-kinase proteins, antioxidant related proteins and some growth-mediating proteins. The variations among gene expressions were validated by qRT-PCR in the two rice varieties. Previous studies indicated that cytoplasmic male sterile lines IR69625A, and G46A had general combining ability for panicle weight and grain yield per plant, while the varieties Giza 179 and Giza 178 had general combining ability for grain yield per plant and seed set percentage. In addition, with regard to the variety Giza 179 which have allelopathic potential and restoring fertility genes, so we recommend by using the variety Giza 179 as parental line with the used cytoplasmic male sterile line to produce hybrid rice have highly yield and strong allelopathy.
Introduction
Weeds are the major biological constraint in crop production this problem has traditionally been solved by hand-weeding. Presently, access to herbicides is helping in decreasing the weed problem. However, increasing farm labour costs and environmental concerns about pesticides usage make it increasingly important to find alternative and sustainable weed management methods. One ecological strategy of weed control, allelopathic activity, has drawn increased attention. Allelopathy is the inhibition of growth of a plant due to biomolecules released by another [1]. The biomolecules are called allelochemicals and are produced by some plants as secondary metabolites. When the allelochemicals are released into the environment, they inhibit the development of neighboring plants. Allelopathic compounds may be released into the environment from plants by means of root exudation, leaching, volatilization and decomposition of plant residues in the soil.
Allelopathy of rice has been shown to be induced by jasmonic and salicylic acids, which are important signaling molecules regulating inducible defense genes against the pathogen infection. Potential allelochemical isolated from root exudates of the rice cultivar Giza 179 is momilactone B. Momilactone B inhibits the growth of typical rice weeds like barnyardgrass and Echinochloa colonum at concentrations greater than 1μmol/L [2]. Rice plants secrete momilactone B from the roots into the rhizosphere over their entire life cycle. The secretion of momilactone B was also confirmed for other rice cultivars. These observations suggest that rice allelopathy may be primarily dependant on the secretion of momilactone B.
In recent years, rice allelopathy has been extensively studied as it is considered as one of the environment- friendly alternative approaches in weed control [3]. Evidences in earlier studies have indicated that rice allelopathy is a quantitative trait, which is mediated by both genetic effects and environmental conditions [4]. The genetic control of allelopathy in rice has been assessed. Studies have shown that rice allelopathy is an inducible trait, which could be mediated by both biotic and a biotic stress conditions such as nutrient starvation and higher accompanying weed densities [5]. Hence, application of plant activators to induce resistance or increase allelopathic potential has become a new strategy for weed control and is expected to be an alternative for effective and sustainable management. However, crucial questions remain unanswered. Investigation of plant activators (e.g., exogenous SA) and their relationship with induced allelopathy would provide valuable information for the understanding of rice plant defense responses and therefore would stimulate discovery of useful resources from allelopathic rice crops and facilitate the development of effective weed control measures. Our work on rice (Oryza. sativa L.) showed that salicylic acid stress enhanced phytotoxic secondary metabolites in both plant tissues and culture solution.
The allelopathic potential of rice was closely related to the types and concentrations of phenolic compounds especially under stress conditions, in which the activities of related enzymes involved in the phenyl propanoid metabolic pathway in allelopathic rice was significantly increased [4]. This has been considered to be related to salicylic acid (SA)-mediated signal transduction pathway [6]. Previous studies have indicated that SA is an immanent compound of phenol in plants and is involved in many physiological reactions as well as signal transduction that trigger off cascade reactions functioning in the induction of systemic acquired resistance (SAR). It was also shown that exogenous SA could induce the accumulation of phenylalanine ammonia-lyase (PAL) mRNA and the synthesis of new PAL protein, resulting in increased enzyme PAL activity in crop plants against pest and weed infection [7].
In this communication, we present a study on the effect of exogenous SA on rice varieties and demonstrate the potential of induced allelopathy in suppression of the target weed. The molecular mechanisms underlying the physiological alterations of rice roots in response to SA treatment will exam and analyze using suppression subtractive hybridization (SSH) and real-time fluorescent quantitative polymerase chain reaction (qRT-PCR) methods. The present investigation was undertaken to study the following objectives; (I) investigation of different gene expression patterns in allelopathic and non-allelopathic rice varieties treated with salicylic acid in rice/weed mixtures, (II) avoid increase the weed seed bank in the soil during field screening and increase the selection efficiency for allopathic activity in rice breeding, (III) increase the yield productivity and improvement the quality of the rice varieties, (IV) decrease both of water usage rate and herbicides during rice season.
Materials and Methods
Plant Materials
This experiment was conducted at experimental farm of Rice Research and Training Center (RRTC), Sakha, Egypt during season 2016 and 2017 and School of Plant, Environmental, & Soil Sciences, Louisiana State University, LA, USA, during season 2018. Nineteen genotypes using a line x tester model, which including four CMS lines, three testers and twelve crosses were used to study the effect of salicylic acid as inducer to allelopathic activity. The experimental design was RCBD in three replicates. Observations were taken on five random plants from each plot. The crop was raised following the package of recommendation practices for summer season at the research farm of RRTC.
Experiment 1: Rice Allelopathy induced by Exogenous SA
Seeds of Rice varieties Giza 179 high allelopathic potential and non-allelopathic rice Sakha 103 as well as barnyardgrass (E. crus-galli L.) will germinate and then sowing on seedling trays, respectively. When the seedlings of rice and barnyardgrass reached 3-leaf and 2- leaf stages, 50 uniform seedlings of each material were selected, transplant into holes spaced at 5 x 6 cm2 in a Styrofoam plate, and a cotton plug was inserting into each hole to stabilize the seedlings. The Styrofoam plate was float on a pot fill with 10 L Hoagland nutrient solution (normal nutrient condition) according to Kim SY, et al., [8]. Three SA treatments were set at the concentrations of 0.1, 0.2, and 0.3 mM. The rice seedlings were sprayed with 10 ml of each SA solution in triplicate, respectively in the mixture of rice/weed and the control was sprayed with the same volume of distilled water.
After treated for 3 days, the roots of 10 rice seedlings and five accompanying barnyardgrass seedlings were sampled from each treatment and the control for biochemical analysis. The roots were rinsed for 10s in distilled water and then the root physiological activity in terms of the assimilated amounts of methylene blue per unit of time was analyzed following the method described by Xiong J, et al., [4]. At 7d after treatment with SA, the rest of barnyardgrass (five plants) in the mixture of rice/weed were sampled and the harvest tissues were snap-kill in an oven at 105C for 20min and then dried at 70C until constant weight. The dry weight (DW) of barnyardgrass was used to determine the inhibition rates (IRs) above control calculated as: IRs = (control - treatment)/control x 100%, which indicates inhibition when > 0 and promotion when < 0 [3]. All data were subjected to analysis of variance using the Statistical Analysis System Program (SPSS). Each value is expressed as the mean of three replicates.
Experiment 2: Molecular Responses to the Induction of Exogenous SA
The seedlings of allelopathic rice varieties as well as the target weed barnyardgrass were prepared as described in experiment 1. After 7 days of recovery in the Hoagland nutrient solution, 30 rice seedlings and 10 weed seedlings were transplant into a Styrofoam plate to form a hydroponic rice-weed mixture with the barnyardgrass seedlings as the target plants (receptor plants) in the center, surrounded by the rice seedlings. The root tissues of the rice seedlings were randomly sampled at 72 h after sprayed with 10 ml of 0.2 mM exogenous SA (treatment) or distilled water (control). After harvested, the samples were immediately frozen in liquid nitrogen and stored at - 80C prior to SSH and qRT-PCR analyzes.
Construction of Subtractive Library
Suppression subtractive hybridization (SSH) according to was employ to construct a subtractive library. Total RNA was extract from the roots of SA- treated rice and its control using the Trizol method (Invitrogen) [9]. Genomic DNA was degrading using DNase I (TaKaRa). cDNA was synthesize using SMART PCR cDNA Synthesis Kit (Clontech). SSH was perform using the PCR-Select cDNA Subtraction Kit (Clontech). After two hybridizations, a nested PCR was used to selectively amplify differentially expressed fragments. PCR products were purifying with E.Z.N.A. Cycle-Pure Kit (OMEGA) and insert into pMD18-T Vector (Takara) and then transform into E. Coli DH5α cells. The transformed cells were plate on 60μg ml-1 ampicillin containing LB agar plates, which were overlaid with 10 mM X-Gal and 50 mg ml-1 IPTG. After incubation overnight at 37 C, white colonies (putative positive clones) were pick and transfer into 1.5 ml tubes containing 500 μl LB liquid medium (AMP+).
Screening and Identification of Clones
Every clone was amplified by PCR (primed with primer 1 and primer 2R in the Clontech PCR-select cDNA subtraction Kit). The clones larger than 300 bp will blot onto Hybond-N+ nylon membrane (Amersham). Tester cDNA and control cDNA was used as forward and reverse probes, respectively to hybridize with the selected clones. Reverse northern blot will performer using the DIG-High Prime DNA Labeling and Detection Starter Kit (Roche). The hybridization membranes were scanned by ArtixScan 1010 (Microtek).
Sequence Analysis
The samples which select sent to School of Plant, Environmental, & Soil Sciences, Louisiana State University, LA, USA, for sequencing. DNA sequences were analyzed at NCBI (the National Center for Biotechnology Information, USA, www.ncbi.nlm.nih.gov) using Blastn and Blastx.
Real-Time Fluorescent Quantitative Polymerase Chain Reaction (qRT-PCR)
qRT-PCR was used to validate the results from SSH analysis and to determine expressional differences of the related genes rice varities. cDNAs was synthesized from equal amounts of total RNA isolate from the roots of allelopathic rice and its counterpart as described above using the ExScript RT Kit (Takara), and the reaction mixture was dilute many times for subsequent PCR with gene specific primers. The qRT-PCR was performing according to the method of SYBR Green I. SYBR Premix Ex Taq (Takara) will used.
Statistic Analysis
All collected data were subjected to statistical analysis using ANOVA as described by ElShamey EAZ, et al., [10]. All statistical analyses were performed using analysis of variance technique by means of "MSTAT" computer soft ware package and “Quantitative genetic analysis”.
Results and Discussions
Allelopathic Characters
Distinct differences in allelopathic potential were observed among the tested rice genotypes, ranging between 25-84% as shown in Table 1, and Giza 179 showed allelopathic activity 85% on Echinochloa crus- galli during 2016 and 2017 season. The parents were crossed with their CMS lines and produced twelve crosses with weed control ranging between 32-87% as shown in Table 2. ElShamey (2016) presented that Rikuto Norin22 and Giza 179 provided that at least 87 and 85% weed control against barnyardgrass in field experiment at RRTC, Egypt. On the other hand, among the crosses, there were some crosses had higher percentage for allelopathic activity as follow; IR69625A × Giza 179, and G46A × Giza 179 and their values were 87 and 86 %, respectively.
For radial area character, it has been found that the parent Giza 179 had higher values among all parents used during 2016 and 2017 seasons as shown in Table 1. This parent was used to be crossed with their CMS lines and produced crosses with values ranging between 11.83– 6.33 cm2, and the high desirable value which was expressed in IR69625A × Giza 179 (11.83 cm2) and G46A × Giza 179 (11.50 cm2) during 2015 and 2016 seasons as shown in Table 2.
These results referred to highly genetic effect which controlled these traits since the environmental effect had no significance. In a similar study, found that the cultivars; Rikuto norine 22, Giza 182 and Vener 1A provided at least 70% weed control in transplanted rice against E. crus- galli under Egypt conditions [11].
| No. | Parents | Radial area (cm2) | Weed control % | ||||||||
| 1 | Giza 179 | 11.37 | 85 | ||||||||
| 2 | Giza 178 | 9.33 | 44 | ||||||||
| 3 | Sakha 103 | 2.3 | 25 | ||||||||
| 4 | IR 69625 A | 5.33 | 39 | ||||||||
| 5 | IR 70368 A | 6.17 | 46 | ||||||||
| 6 | G46A | 5.5 | 35 | ||||||||
| 7 | K17A | 6 | 37 |
Table 1: Mean performance of radial area and weed control for the parental lines.
| Radial area | c | Weed | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| No | . | Crosses | |||||||||
| (cm2) | ontrol % | ||||||||||
| 1 | IR69625A × Giza 179 | 11.83 | 87 | ||||||||
| 2 | IR69625A × Giza 178 | 8.17 | 65 | ||||||||
| 3 | IR69625A × Sakha 103 | 6.33 | 35 | ||||||||
| 4 | IR70368A × Giza 179 | 11.33 | 83 | ||||||||
| 5 | IR70368A × Giza 178 | 6.33 | 65 | ||||||||
| 6 | IR70368A × Sakha 103 | 7 | 30 | ||||||||
| 7 | G46A x Giza 179 | 11.5 | 86 | ||||||||
| 8 | G46A x Giza 178 | 8 | 60 | ||||||||
| 9 | G46A x Sakha 103 | 6.5 | 37 | ||||||||
| 10 | K17A x Giza 179 | 11.33 | 85 | ||||||||
| 11 | K17A x Giza 178 | 8.5 | 63 | ||||||||
| 12 | K17A x Sakha 103 | 6.83 | 37 |
Table 2: Mean performance of radial area and weed control for the obtained crosses.
Morphological Characters
A wide range for morphological characters of the parental lines with their crosses was recorded, the data summarized in Table 3 show that the mean values of the three testers and the two CMS lines were (86.71 cm; 93.14 days; 37.11 cm2 and 30.43 ô) for plant height, days to heading, flag leaf area and flag leaf angle, respectively. But, the mean values for the F1 hybrids were (97.76 cm; 92.48 days; 36.52 cm2 and 24.81 ô) for plant height, days to heading, flag leaf area and flag leaf angle, respectively as shown in Table 3.
For plant height, the results in Tables 3 and 4 revealed that the most desirable mean values towards dwarf were found for the parents IR70368A (71.33 cm) and IR69625A (76 cm) in addition to, the crosses and K17A × Sakha103 and K17A × Giza 179 which gave the lowest values for plant height. The selection of parental lines with appropriate plant height and non–loading characters is important for high yield potential hybrids, particularly interspecific hybrids [12].
| No. | Parents | Plant height | Days to heading | Flag leaf area | Flag leaf angle | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Giza 179 | 96 | 90 | 46.42 | 32 | |||||||||||
| 2 | Giza 178 | 111 | 98 | 44 | 23.33 | |||||||||||
| 3 | Sakha 103 | 87.33 | 93 | 32.99 | 35 | |||||||||||
| 4 | IR 69625 A | 76 | 100 | 33.33 | 35 | |||||||||||
| 5 | IR 70368 A | 71.33 | 99 | 44.67 | 26.33 | |||||||||||
| 6 | G46A | 83.33 | 87 | 31 | 28.33 | |||||||||||
| 7 | K17A | 82 | 85 | 27.33 | 33 | |||||||||||
| Mean | 86.71 | 93.14 | 37.11 | 30.43 |
Table 3: Mean performance of morphological characters for the parental lines.
With respect to days to heading, the most desirable mean values towards the earliness were obtained from the parents; Giza179 (90 days) and Sakha103 (93 days) and the crosses; IR70368A × Giza179, IR69625A × Giza 179 and IR70368A × Giza 178 as shown in Tables 3 and 4, whereas the selection for this trait should be done with the plants which have the lowest days to heading. For flag leaf area, the parents; Giza179 and IR70368A and the Regarding to flag leaf angle, the testers; IR69625A and Sakha 103 and the crosses, IR69625A × Giza 179 and IR58025A×Giza182 gave the desirable mean values, indicating that IR69625A was a good donor for the desirable flag leaf angle.
crosses IR69625A × Giza 179 gave the desirable mean values.
| No. | Crosses | Plant height | Days to heading | Flag leaf area | Flag leaf angle | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | IR69625A × Giza 179 | 101 | 93 | 44.63 | 35 | |||||||||||
| 2 | IR69625A × Giza 178 | 113.33 | 97 | 25.73 | 16.67 | |||||||||||
| 3 | IR69625A × Sakha 103 | 97.67 | 98.67 | 34.25 | 26.67 | |||||||||||
| 4 | IR70368A × Giza 179 | 101 | 92 | 34.28 | 24.33 | |||||||||||
| 5 | IR70368A × Giza 178 | 103.33 | 93.33 | 26.51 | 21.67 | |||||||||||
| 6 | IR70368A × Sakha 103 | 94.33 | 98 | 31.48 | 25 | |||||||||||
| 7 | G46A x Giza 179 | 97.33 | 89.5 | 61.7 | 25.7 | |||||||||||
| 8 | G46A x Giza 178 | 96 | 91 | 34.8 | 25 | |||||||||||
| 9 | G46A x Sakha 103 | 93.33 | 95.87 | 33.33 | 24.33 | |||||||||||
| 10 | K17A x Giza 179 | 91 | 79.85 | 39.51 | 22.3 | |||||||||||
| 11 | K17A x Giza 178 | 94.5 | 85.2 | 39.56 | 25.5 | |||||||||||
| 12 | K17A x Sakha 103 | 90.33 | 96.33 | 32.5 | 25.5 | |||||||||||
| Mean | 97.76 | 92.48 | 36.52 | 24.81 |
Table 4: Mean performance of F1 crosses for some morphological characters.
Yield and its Component Characters
The mean performance of yield and its component characters i.e., number of panicles per plant, panicle length, panicle weight, grain yield per plant, number of grains per panicle, 1000-grain weight and seed set (%) for the parental lines are presented in Table 5.
No. of grains/ panicle
- No.
- Parents
- No. of panicles / plant
- Panicle
- Panicle
- Weight
- Grain
- Yield/plant length
- 1
- Giza 179
- 15
- 23
- 4.83
- 30.4
- 167
- 21.43
- 92.5
- 2
- Giza 178
- 22
- 23.33
- 4.56
- 37.67
- 155
- 21
- 96.57
- 3
- Sakha 103
- 13.33
- 18
- 3.74
- 25.33
- 126
- 24.27
- 81
- 4
- IR 69625 A
- 16
- 21
- 1.53
- 10
- 124
- 22
- 32
- 5
- IR 70368 A
- 17
- 23.17
- 2.3
- 17
- 128
- 22.67
- 31
- 6
- G46A
- 16
- 21.7
- 3.1
- 19
- 125
- 26.6
- 32
- 7
- K17A
- 18
- 21.5
- 3.95
- 16
- 123
- 27.1
- 29
- Mean
- 16.76
- 21.67
- 3.43
- 22.2
- 135.43
- 23.58
- 56.3
Table 5: Mean performance of yield and its component characters for the parental lines.
A wide range for yield and its component traits of the parental lines with their crosses was recorded, the data summarized in Table 5 show that the mean values of the three CMS lines and the ten testers were (16.67, 21.67, 3.43, 22.20, 135.43, 23.58 and 56.30) for number of panicles per plant, panicle length, panicle weight, grain yield per plant, number of grains per panicle, 1000-grain weight and seed set (%), respectively. The decrease of the mean values for grain yield per plant referred to the decrease of out-crossing to the CMS lines. But, the mean values performance of F1 hybrids for these characters were (18.28, 22.14, 4.14, 41.31, 132.25, 25.79 and 79.48), respectively, as shown in Table 6.
From the results summarized in Tables 5 and 6 it could conducted that for number of panicles per plant character, the highest mean values were obtained by the two parent Giza178 (22.00) and the cross IR70368A×Giza 179 which gave mean values of 26.00 as shown in Table 5.
The desirable mean values for panicle length were recorded by the three parents; Giza 178, IR70368A and Giza 179 with values of (23.33, 23.17 and 23.00 cm), respectively (Table 5) and the crosses; IR69625A × Giza 179 and IR69625A×Giza 178, gave also the desirable mean values (25.00 for first cross and 23.33 for the second cross) as shown in Table 6.
While, for panicle weight, the parents Giza 179 and No. Crosses No. of panicles Panicle / plant length set 1 IR69625A × Giza 179 14 25 3.51 45.6 115 26.1 94.87 2 IR69625A × Giza 178 18.33 23.33 4.53 55.67 173 27 98.8 3 IR69625A × Sakha 103 16.67 19.33 2.67 16.33 123 24.33 48.43 4 IR70368A × Giza 179 26 21.5 3.62 59.4 145 27.5 79.27 5 IR70368A × Giza 178 12 23 4.6 65 152 25 98.37 6 IR70368A × Sakha 103 14.33 18.5 2.37 15.33 119 25.33 46.3 Giza178 gave the desirable mean values (4.83 and 4.56), respectively in Table 5, and the crosses; IR70368A × Giza 178 and IR69625A × Giza 178, gave the desirable mean values (4.60 and 4.53), respectively (Table 6).
For grain yield per plant character, the parents Giza 178 and Giza 179 which gave (37.67 and 30.40 gm), respectively and the two crosses IR70368A × Giza179 and IR70368A × Giza 178 which gave (65.00 and 59.40), respectively recorded the highest mean values for this character. While, the tester parents, Giza 179 and Giza 178 with mean values of (167.00 and 155.00), respectively and the two crosses; IR69625A×Giza178 and IR70368A×Giza 178 with mean values of (173.00 and 152.00), respectively recorded the highest mean values for the no. of grains per panicle character.
In addition, with regard to the 1000-grain weight character, the desirable mean values were recorded for the tester lines, Sakha 103 and Giza 179 which gave (24.27 and 21.43 gm), respectively and the crosses; IR70368A × Giza 179 (27.50) and IR69625A x Giza 178.
Also, it could be noticed that the desirable mean values for seed set % character were recorded for the tester Giza 178 which gave (96.57) and the cross IR69625A×Giza178 which gave (98.80). This hybrid may be considered as ideal sources for varieties improvement through pedigree breeding or biparental meeting.
Panicle Weight
Grain Yield/
No. of grains/
1000-grain Seed
plant panicle weight
| 7 | G46A x Giza 179 | 21.9 | 25.6 | 7.1 | 64.9 | 123 | 26.1 | 98 | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 8 | G46A x Giza 178 | 20.9 | 24.8 | 5.5 | 48.9 | 137 | 24.8 | 96 | ||||||||||||||
| 9 | G46A x Sakha 103 | 17 | 18.87 | 2.87 | 17.87 | 119 | 24.5 | 52 | ||||||||||||||
| 10 | K17A x Giza 179 | 19.3 | 23.7 | 5.3 | 48 | 122 | 28.2 | 96.8 | ||||||||||||||
| 11 | K17A x Giza 178 | 21 | 23.1 | 4.65 | 42.9 | 139 | 25.9 | 95 | ||||||||||||||
| 12 | K17A x Sakha 103 | 17.87 | 19 | 3 | 15.87 | 120 | 24.67 | 4 | 9.87 | |||||||||||||
| Mean | 18.28 | 22.14 | 4.14 | 41.31 | 132.25 | 25.79 | 7 | 9.48 | ||||||||||||||
Table 6: Mean performance of yield and its component characters for the obtained crosses.
Analysis of Variance
The analysis of variance (Table 7) revealed highly significant differences among the 19 genotypes (12 hybrids, 3 testers, and 4 CMS lines) tested for all studied characters. The parental lines and the hybrids showed highly significant differences for all characters. These results indicate that genotypic differences between entries are present. Mean square values of parents and crosses were found to be highly significant for all studied characters. These results could be used as an indication to average heterosis overall crosses and therefore could be used through hybrid breeding technology to improve such characters. Parents V.S crosses mean square indicated that average heterosis was significant in all crosses for all characters, the variances in all studied traits differed significantly with different combinations, and this is agreement with ElShamey EAZ, et al., [13].
- Flag
- Flag
- No. of panicles/
- Grain
- Yield/
- Plants
- No. of grains/ panicle
- Source of variance d.f. Radical
- Weed control
- Plant height
- Days of heading panicle panicle
- 1000 grain leaf area leaf angle area length weight plants
- Reps
- 2
- 0.16
- 0.002
- 0.635 n.s. 1.73 n.s.0.362
- 2.28 n.s.
- 0.09 n.s. 1.45 n.s. 0.01 n.s0.64 n.s 4.73 n.s. 0.04 n.s.
- 0.001 n.s. n.s. n.s.
- Entries
- 18 27.12 0.167 519.91 49.84 131.36
- 104.26
- **
- 43.94**
- 17.87 4.48 1362.351590.88 10.95**
- 0.225**
- **
- Crosses
- 11 2.28* 0.018* 14.39*
- 2.77* 13.28* 9.77*
- 2.87*
- 1.87*
- 0.44* 179.97* 645.17*
- 87.99**
- 0.023*
- Parents
- 6 37.19 0.148 732.77 87.9 93.9181.65
- 53**
- 18.07 5.54 754.86 1439.4
- 5.81**
- 0.334**
- P*V.C.
- 1 66.01 0.880 2174.06
- **
- 110.95** 851.52
- 645.09
- **
- 191.09 75.10 20.11 9682.263128.26 43.13
- 1.072**
- **
- Error
- 18 4.48
- 0.001
- 2.073
- 1.854
- 1.93
- 1.75
- 2.18
- 1.91
- 0.04*
- 2.07
- 7.85
- 0.04
- 0.002
Table 7: Mean square estimates of the ordinary analysis for allelopathic characters, morphological characters, and yield
Allelopathic Effect in Rice induced by Salicylic Acid
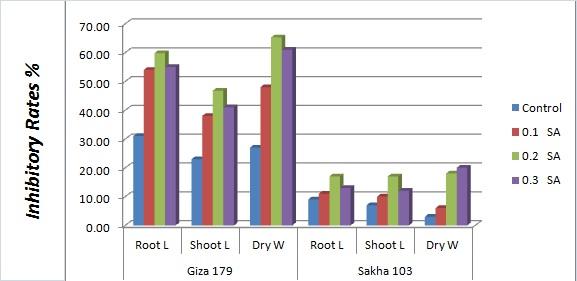
Allelopathic rice varieties Giza 179 showed higher inhibitory effect on the barnyardgrass than Sakha 103 after the two rice varieties were treated with different concentrations of salicylic acid; 0.1, 0.2, and 0.3 mM/L. The inhibitory rates (IRs) was enhanced with the increase of SA concentration and reached a peak at 0.2 mM with 59.81% for suppression of root length, 46.76% for suppression shoot length, and 65.28% for suppression to dry weight of barnyardgrass (Figure 1). However, a slight decreasing tendency of the inhibitory rates was found when the treated concentration of salicylic acid was increased to 0.3 mM. A similar tendency was found for the non allelopathic rice Sakha 103, but its inhibitory effect was always lower than Giza 179 in all of the SA treatments. This result indicated that salicylic acid treatment with an appropriate dosage could significantly enhance the allelopathic effect of rice on barnyardgrass, especially in allelopathic rice varieties Giza 179.
Induced Gene Expression in Allelopathic Rice
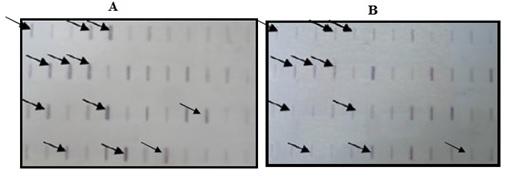
A library of cDNA was constructed by Suppression Subtractive Hybridization (SSH) for determination of expressed genes in allelopathic rice Giza 179 in response to salicylic acid treatment. A total of 728 clones were obtained from the forward subtractive library, 96 of which were selected for reverse northern blot after PCR (Figures 2A & B). The expressions of 14 clones (genes) were found to be up-regulated. These clones were sequenced, and the cDNA sequences were searched against standard databases (www.ncbi.nlm.nih.gov). According to nucleic acid homologies and encoded protein sequences of the rice (Oryza sativa L.) database, clones were successfully assigned. The putative functions of the assigned expressed sequence tags (ESTs) are given in Table 1. Clones 171 and 459 showed the highest homology to a putative receptor-like protein kinase from rice (Oryza sativa L.). Clones 163 and 617 encode the same ubiquitin carrier protein, and clones 557, 695 code for a serine/threonine protein kinase-like protein and BAG domain containing protein-like protein, all of which are involved in signal transduction.
Clone 265 encodes a putative glutathione S- transferase functioning in detoxification in rice plants and clones 670, 438, 512, 587 and 316 had the highest homology to a cellulose synthase-4, a putative acetyltransferase, a putative cinnamoyl-CoA reductase, a putative endosperm specific protein SC3, and peptidylprolyl cis-trans isomerase from Oryza sativa, respectively, which are related with substance metabolism. Clones 593 and 507 had homology to a putative peroxidase (POD) and phenylalanine ammonia- lyase from Oryza sativa, respectively, which are related with de novo biosynthesis of phenolic allelochemicals involved in plant chemical defence against salicylic acid treatment. These identified genes can be categorized into five groups according to their function; signal transduction, growth-mediated transcription regulation, secondary metabolism, cell defence, protein synthesis and its degradation.
Validation of Differentially Expressed Genes by qRT-PCR
Differential expression of two genes by qRT-PCR gene expression of both PAL and POD, which are involved in de novo allelochemical synthesis was increased in Giza 179 and Sakha103 when exposed to salicylic acid treatment. The relative expression levels of the two genes in Giza 179, however, were higher than those in Sakha103, showing that the relative transcript abundance of PAL in root of Giza 179 increased by 11.38 times, while in Sakha103, they were increased by 1.15 times after three days after treatment with 0.2 SA, respectively. The same was true in the expression of P450 gene for the two rice varieties as shown in Fig. 3. This finding further confirms the findings of SSH analysis (Table 1).
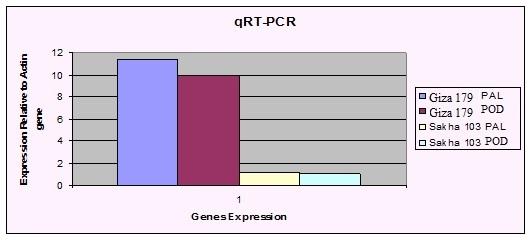
Figure 3: SYBR-Green RT-PCR Analysis of Phenolic- based allelochemicals Pathway Gene Expression in Giza 179 and Sakha103. Total RNA was reverse transcribed, and aliquots were amplified using primer pairs specific for PAL and POD. The present results confirmed that SA could serve as an elicitor to induce higher allelopathic potential of donor rice to suppress the target weed plants. This inducible effect is respondent to the dose of SA treatment. The allelopathic inhibitory effect of Giza 179 on barnyardgrass was significantly increased after the rice varieties was treated with SA. The activities of protective enzymes PAL and POD as well as root physiological capability were all enhanced in Giza 179, implying that the plant chemical defence in rice has taken effect in the suppression of the
- target weed. Here, we present an update on the effects of
- SA treatment on the expression of genes involved in rice plant chemical defence response using the combined approach of SSH and qRT-PCR. Our study showed that many genes in the rice varieties can be significantly upregulated by exogenous SA treatment.
- Clone Length
- (bp) Accession no.
- Best homologue database score E-value/Sequence identify (%)
- Source
- 171
- 324
- AC131374
- Putative receptor-like protein kinase
- 278
- 2e-73/100
- Oryza sativa
- 187
- 382
- AA134132
- Putative acetyltransferase
- 229
- 8e-05/92
- Oryza sativa
- 245
- 195
- ABR25322
- Phenylalanine ammonia-lyase
- 176
- 2e-73/100
- Oryza sativa
- 311
- 221
- AP003734
- Putative cinnamoyle-CoA reductase
- 231
- 3e-34/100
- Oryza sativa
- 418
- 219
- AY224431 Serine/threonine protein kinase-like protein 356
- 2e-45/97
- Oryza sativa
- 434
- 297
- AP005578
- Putative endosperm specific protein SC3
- 143
- 2e-64/100
- Oryza sativa
- 519
- 313
- AP003252
- BAG domain containing protein-like
- 143
- 4e-105/100
- Oryza sativa
- 543
- 349
- CM000126
- Ubiquitin carrier protein
- 76
- 0/97
- Oryza sativa
- 567
- 245
- AP000263
- Putative peroxidase
- 45
- 4e-127/98
- Oryza sativa
- 590
- 314
- CM000126
- Ubiquitin carrier protein
- 77
- 5e-92/98
- Oryza sativa
- 598
- 221
- AC131374
- Putative receptor-like protein kinase
- 111
- 3e-90/100
- Oryza sativa
- 601
- 248
- AP000528
- Cellulose synthase-4
- 196
- 2e-54/100
- Oryza sativa
- 623
- 291
- AC092236
- Putative glutathione S-transferase
- 111
- 8e-125/99
- Oryza sativa
- 667
- 288
- CM000126
- Ubiquitin carrier protein
- 166
- 5e-154/98
- Oryza sativa
Table 8: Sequence similarity comparison display fragments genomic library of the root in allelopathic rice Giza 179 under
Both defense-related proteins and the key enzymes such as PAL, CCR, GST, and POD, etc., which are associated with phenylpropanoid metabolism and plant defense, showed significantly positive expression in allelopathic rice Giza 179. It has been reported that phenylalanine ammonia-lyase (PAL), which can be induced by biotic and a biotic factors, is a key enzyme of plant metabolism catalyzing the first reaction in the phenylpropanoid biosynthesis from L-phenylalanine to a wide variety of natural products based on the phenylpropane skeleton [14, 15]. More recently, work by has suggested that under conditions of decreased nitrogen availability, plant phenolic content and PAL activity might increase because of decreased demand for proteins involved in growth, and ammonium ions released by PAL could be assimilated to increase nitrogen cycling [16]. The result also indicated that the gene coding for CCR, which is the entry-point enzyme of the lignin specific biosynthetic pathway, was up-regulated in this SA-mediated process. Previous studies showed that the product lignin released from the donor plants functioned as specific inhibitors of the detoxifying enzymes (e.g., P450s) present in the receiver plant (e.g., barnyargrass) to block the ability of modifying toxic agents [17]. As a result, chemical defence could take effect at a lower dosage of phytotoxin. The present result suggested that the SA mediated cascade reaction in rice to defend against barnyardgrass attack might be similar to the responses to nutrient deficiency through the regulation of carbon-nutrient balance. This in turn results in the accumulation of phenolics and more phenolic allelochemicals being released into external environment (culture solution) from the roots to depress the target weed growth including the inhibition of protective enzyme activity and root uptake ability, which was consistent with the previous studies [18]. This raises the question as to why the auto-toxicity does not occur during this process.
Our present work elucidated that the donor plants possess a detoxifying system and that the up-regulation of the enzyme might play an important role in this process. It has been reported that this enzyme, which could be activated by SA, was associated with xenobiotic detoxification, limiting oxidative damage, and other stress responses in plants [19]. The present work suggested that the variety Giza179 is strong allelopathy, and from obvious studies we found the variety Giza179 as restorer line [15]. So, we recommend by using variety Giza 179 as parental line to produce hybrid rice IR69625A x Giza179 and IR70368A x Giza 179 which have highly yield and strong allelopathy.
Acknowledgements
This project was supported by Ministry of Higher Education & Scientific Research, Science & Technology Development Fund, Research Grant number 23152 to Essam Adel Zaki ElShamey, Agricultural Research Center, Field Crops Research Institute, Rice Research Department.
References
-
He HQ, Shen LH, Xiong J, Jia XL, Lin WX, et al. (2004) Conditional genetic effect of allelopathy in rice (_Oryza_ _sativa_ L.) under different environmental conditions. Plant Growth Regul 44(3): 211-218.
-
ElShamey EAZ (2016) Genetical studies on allelopathic activity and yield component in some lines of hybrid rice against barnyardgrass weed under Nitrogen stress. Asian Journal of Plant Science and Research 6(4): 30-36.
-
Lin WX, Kim KU, Shin DH (2000) Allelopathic potential in rice (_Oryza sativa_ L.) and its modes of action on barnyardgrass (Echinochloa crusgalli L.). Allelopath J 7(2): 215-224.
-
Xiong J, Lin WX, Zhou JJ, Wu MH, Chen XX, et al. (2005) Studies on biointerference between barnyardgrass and rice accessions at different nitrogen regimes. _In_: Harper JDI, An M, Wu H, Kent JH Proceedings fourth world congress on allelopathy, Charles Sturt University, Wagga Wagga, NSW, Australia, pp: 501-504.
-
Shen LH, Lin WX (2007) Effects of phosphorus levels on allelopathic potential of rice co-cultured with barnyardgrass. Allelopath J 19: 23-32.
-
Shen L, Xiong J, Lin WX (2008) Rice allelopathy research in china. Allelopathy in sustainable agriculture and forestry pp: 215-233.
-
Bi HH, Zeng RS, Su LM, An M, Luo SM (2007) Rice allelopathy induced by methyl jasmonate and methyl salicylate. J Chem Ecol 33(5): 1089-1103.
-
Kim SY, Madrid AV, Park ST, Yang SJ, Olofsdotter M (2005) Evaluation of rice allelopathy in hydroponics. Weed Res 45(1): 74-79.
-
Diatchenko L, Lau YFC, Campbell AP, Chenchik A, Moqadam F, et al. (1996) Suppression subtractive hybridization: a method for generating differentially regulated or tissue-specific cDNA probes and libraries. Proc Natl Acad Sci USA 93(12): 6025-6030.
-
Gomez KA, Gomez AA (1983) Statistical Procedures for Agricultural Research. (2nd Edn) Jhon Wiley and Sons, New Yourk, USA pp: 680.
-
ElShamey EAZ (2012) Genetical studies on restoring ability and allelopathic activity on some lines of hybrid rice. Ph.D. Thesis, Fac Agric Tanta Univ Egypt.
-
Ikehashi H, Zou JS, Moon HP, Maruyama A (1994) Wide compatibility gene(s) and indica-japonica heterosis in rice for temperate countries. In: Virmani SS, Hybrid Rice technology: New Development and Future Prospects, IRRI, Manila, Philippines pp: 21-31.
-
Sharma RK, Mani SC (2012) Combining ability studies for grain yield and other associated characters in basmati rice (_Oryza sativa_ L.). Crop Improvement 28: 236.
-
ElShamey EAZ, ElSayed MAA, ElGamal WH (2015) Genetical analysis for allelopathic activity in some rice varities. Egypt J Plant Breed 19(3): 125-137.
-
Bob BB, Wilhelm G, Russell LJ (2000) Biochemistry & molecular biology of plants. American Society of Plant Physiologists, Maryland.
-
ElShamey EAZ, ElSayed MAA, ElGamal WH (2016) Genetical studies on the relationship of isozymes with allelopathy in hybrid rice. KMITL Sci Tech J 16(2).
-
ElDenary IM, ElShamey EAZ (2014) Analysis of gene expressions associated with increased allelopathy in rice (_Oryza sativa_ L.) under low nitrogen treatment. Egypt J Genet Cytol 43(1): 113-131.
-
Song BQ, Xiong J, Fang CX, Qiu L, Lin RY, et al. (2008) Allelopathic enhancement and differential gene expression in rice under low nitrogen treatment. J Chem Ecol 34(5): 688-695.
-
Gong HB, Jiao YX, Hu WW, Pua EC (2005) Expression of glutathione S-transferase and its role in plant growth and development in vivo and shoot morphogenesis in vitro. Plant Mol Biol 57(1): 53-66.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note