Effect of Tragacanth Gum on Some Protective Mechanisms of Black Cumin (Nigella sativa L.) Under Drought Stress Conditions
The production of active oxygen species increases under drought stress conditions, which means that their removal or deactivation is out of plant power. Therefore, most plants in the face of drought stress require solutions adopted by farm managers. In order to investigate the effect of various irrigation regimes and anti-evapotranspiration effect of different concentrations of tragacanth gum on protective mechanisms of Nigella sativa L., a factorial experiment was conducted in a completely randomized design with three replications in the greenhouse of the Faculty of Agriculture, the University of Kurdistan in 2018. The experimental factors included irrigation at three levels of 100 (full irrigation), 70 (mild drought stress), and 40% (severe drought stress) of field capacity of soil and spraying with tragacanth gum at six concentrations of 0, 1.25, 2.5, 5, 7.5, and 10 g/L. The results showed that increasing the intensity of drought stress (irrigation reduction) resulted in increased levels of H2O2, MDA, SOD, POD, proline, soluble carbohydrates (water and alcohol soluble), and osmotic potential of the plant and reduced grain yield. The effect of different concentrations of tragacanth gum was differed at different levels of irrigation. In full irrigation, the concentration of 1.25 g/L was positive for all studied traits. In mild drought stress, the use of higher concentrations of tragacanth (up to 5 g/L) had the best efficiency, and concentrations greater than 5 g/L had opposite effects on the traits. In severe drought stress, the use of higher concentrations of tragacanth was beneficial and concentrations up to 7.5 g/L improved the studied traits, but concentrations of 10 g/L had a negative effect on these traits. Regarding that the application of appropriate concentrations of tragacanth gum improved the measured traits and increase grain yield under mild and severe drought stress by 11.6% and 28.2%, respectively, it may be introduced as a novel anti-evapotranspiration agent with natural origin and may be useful in areas with drought stress.
Introduction
Due to the positive approach of people towards the use of herbal drugs and their unwillingness to use synthetic drugs, as well as the cost-effectiveness of producing herbal plants, the cultivation of these plants can be considered common in arid and semi-arid regions. Nigella sativa L., one of the forgotten herbal plants, has industrial and medical applications for the treatment of diseases such as bronchitis, rheumatism, high blood pressure, cough, eczema, inflammation, and influenza [1]. Improving the growth and yield of this plant and other medicinal plants, especially in areas with water shortages, is important to meet the medical needs of the world’s population. Ecological studies have shown that climate change and the occurrence of warm and dry periods, caused by it Hoerling M, et al. [2], Hertig E, et al. [3], Naumann G, et al. [4], affect the food and pharmaceutical needs of the growing population [5, 6]. Although semi-arid areas are suitable for the production of medicinal plants (due to the desired amount of solar radiation), optimal production of these plants, like other plants, requires adequate water and irrigation [1]. Medicinal plants have a low 1000-grains weight and are often cultivated when the air is heated and rainfall is limited in the Mediterranean, so drought stress can be considered the most important limiting factor in the production of medicinal plants in the Mediterranean region. The effect of drought stress on the growth and yield of black cumin grain is mainly due to the disruption of metabolic pathways [7]. Drought stress restricts plant photosynthesis and causes changes in chlorophyll content and damage to photosynthetic structures. One of the important reasons why environmental stresses such as drought reduce plant growth and photosynthetic ability is the disturbance in the balance between the production and removal of free oxygen radicals [8]. The consequence of the production and accumulation of free oxygen radicals is the peroxidation of unsaturated fatty acids. The peroxidation of unsaturated fatty acids results in the production of aldehydes, such as Malondialdehyde (MDA), which is usually evaluated as an oxidative stress index [9]. Various types of active oxygen can also attack vital cellular components such as unsaturated fatty acids, proteins, and nucleic acids. These reactions naturally reduce membrane fluidity, ion transfer, enzymatic activity, and protein synthesis, leading to the destruction of nuclear and mitochondrial DNA, and eventually cell death [10].
Under drought stress conditions, high activity of antioxidant enzymes and high content of non-enzymatic compounds are important for plant tolerance to stress, and plants adopt enzymatic and non-enzymatic strategies to cope with oxidative stress [11]. Antioxidant mechanisms such as superoxide dismutase (SOD) and peroxidase (POD) play an essential role in balancing the production and removal of free oxygen radicals [8]. The first mechanism for the removal of active oxygen species is SOD. The SOD enzyme, by producing H2O2, removes anion superoxide (O-
2), but H2O2 production, in turn, is harmful and should be removed from the cell. Hydrogen peroxide is removed in the next step by CAT and POD enzymes, and thus the cell is cleaned from harmful compounds [12]. The accumulation of compatible osmolytes is also a common response of plants to counteract drought stress. Proline and soluble sugars are compatible osmolytes, which increase in different plants under the influence of environmental stresses. Proline is a cell wall compound and an osmoprotectant, which helps to stabilize the membrane under the influence of stress. Proline content increases during drought stress, which can be due to increased proline synthesis, but increasing the amount of proline under stress conditions, in addition to biosynthetic activity, also depends on the cessation of its destructive activity. Proline is an osmoprotectant and plays a role in maintaining the water balance, maintaining the three- dimensional structure of proteins and enzymes, reducing the risks of ROS production, removing hydroxyl radicals, and quenching the singlet oxygen (maintenance of cellular macromolecules such as proteins, lipids, and DNA from the dangers of free radicals), biological membrane stability and protein synthesis mechanism, regulating cellular pH and the ratio of NADP+/NADPH. Proline also acts as a source of carbon and nitrogen storage for plant growth after stress [13]. Proline is also effective in reducing the optical deterrence damage in the thylakoid membrane by removing or reducing the production of singlet oxygen [14].
Carbohydrates play a double role in plant cells. On the one hand, carbohydrates, as an osmotic agent, cause the negative osmotic potential and maintain the turgor and vitality state of the cells, and on the other hand, by supplying the energy and carbon skeleton required for biosynthesis processes, cause the growth and development of the cells [15]. Evaluating the effect of drought stress on the soluble carbohydrate content of Stevia rebaudiana showed a decrease in soluble carbohydrates [16]. Saucedo CCM, et al. [17] investigated the effect of drought stress on bean carbohydrate concentrations. They reported that drought stress at the flowering stage for 6 and 10 days increased glucose levels (78.5% and 196%) and fructose (190% and 233%) in mature leaves. The amount of water consumed by the plants for photosynthesis is less than 5% of the plant’s total water use, and the remainder is sent out by transpiration from the plant. Although transpiration is a necessary process for photosynthesis and growth of plants, depending on the conditions, it may be harmful in some cases, so the use of anti-transpirant can be one of the most effective methods for reducing the amount of water lost through transpiration. The anti-transpirant improves the damaging effects of drought stress on plants due to increased leaf resistance to water loss. Reported that corn yield increased due to the use of anti-transpiration agents. They also stated that anti-transpiration agents increased the relative content of leaf water without disturbing gas exchange. Tragacanth is a natural gum and anionic polysaccharide that is secreted from Asian species of Astragalus plant. Tragacanth plant is found in Turkey, Iraq, Syria, Lebanon, Afghanistan, Pakistan, Russia, and in most parts of Iran, especially in the central area to the west and in the Zagros mountainous regions. Iranian tragacanth has the highest quality among tragacanths produced in these countries. Tragacanth is both a stabilizer and reduces surface tension. It has long been widely used in the food, pharmaceutical, cosmetics, textile, and medical industries.
Climate change created in the world and the intensification of stresses caused by it, especially drought stress in arid and semi-arid regions such as Iran, makes it difficult to produce medicinal plants such as black cumin. Therefore, finding strategies that can reduce the effects of water deficiency on plant and improve growth and yield can be very important. According to the coating property of tragacanth gum, this idea came to the mind of the authors of this paper that this substance could be used as an anti-transpirant coating to reduce water loss through transpiration without reducing plant growth and yield. The aim of this study was to investigate the effect of spraying of various doses of tragacanth gum on the activity of antioxidant enzymes, compatible osmolytes (proline and soluble carbohydrates) and grain yield of black cumin under different irrigation regimes, so that in the presence of positive effects on plant growth and yield in conditions of water deficiency, the tragacanth matter can be introduced as a new anti-transpiration agent with organic origin to agriculture.
Materials and Methods
Experiment Conditions and Treatments
This research was carried out in a factorial experiment in a completely randomized design with three replications in a greenhouse of the Agriculture Faculty of Kurdistan University from May 31 to September 11, 2018. The experimental factors included irrigation at three levels of 100% (full irrigation), 70% (mild drought stress), and 40% (severe drought stress) of the field capacity of the soil, and spraying with tragacanth gum at six concentrations of 0, 1.25, 2.5, 5, 7.5, and 10 g/L. During the experiment, the minimum and maximum greenhouse temperatures were 18 and 33°C, respectively, and the relative humidity was 27%. The required light was supplied with the natural light of the sun, and considering the installation of the sunshade on the greenhouse, the average light intensity was about 400 to 600 µmoles/m2/s. First, plastic pots (30 cm in diameter and 30 cm in height, 706.5 cm area, and 400 g in weight) were filled with 11316 g of soil (Table 1). Each pot reached the field capacity with 1734 g of water. Before planting, according to soil analysis, 25 kg of nitrogen per hectare of urea source, 50 kg/ha of potassium (K2O), and 70 kg/ha of phosphorus (P2O5) were added to the soil. In each pot, 14 seeds of black cumin (200 plants per square meter) were cultivated. Irrigation treatments were applied one week before flowering (in the budding stage) and continued for 40 days (9 stages) and up to 10 days before harvest (physiological maturity).
Prior to applying irrigation treatments, irrigation was performed completely for all treatments. The pots were moved at each irrigation fold, in order to reduce or eliminate the error caused by the location of the pots in the greenhouse. The time of applying irrigation treatments (percentage of field capacity) in pots was determined by the weighing method. Irrigation was carried out at intervals of 5 days. In each irrigation fold, the weight of the pot under full irrigation and without spraying with tragacanth was the criterion, and after 5 days, the amount of water needed to reach the field capacity was determined and consumed.
For 40% and 70% irrigation treatments, 40% and 70% of the water required for full irrigation treatment were used. In pots where their required water was less than the water consumed in full irrigation without spraying, excess water was calculated as stored water to be considered in calculating the water use efficiency. At the same time as applying irrigation treatments, tragacanth extract was sprayed at the beginning of budding. For this purpose, weighed tragacanth values from the rangelands of Kurdistan province (A gummifer) were dissolved in 1 liter of tepid water. After 24 hours, it was mixed with a mixer for 5 minutes to be completely uniform. Spraying of this material was done using a back sprayer (SHARK model) with a constant pressure of 2.4 bar and a volume of 250 liters of water per hectare. In order to prevent splashing of the material into adjacent pots, a nylon curtain was used between the pots at the time of spraying.
Features of Tragacanth
Tragacanth is a high-quality hydrocolloid that is a non- uniform and highly branched hydrophilic carbohydrate in terms of chemical composition. Tragacanth consists of two main components called Tragacantic acid or Basourin and tragacanthin. Basourin accounts for 60-70% of the total gum and is a water-insoluble component that has the ability to swell and form a gel. The other component, tragacanthin, is dissolved in water and leads to the formation of a colloidal solution. Basourin is an acidic component that produces sugars such as D-xylose, L-fucose, D-galacturonic acid, and a small amount of L-Rhamnose through acid hydrolysis. The acid component of this molecule is also related to cations of calcium, magnesium, and potassium. The properties of tragacanth gum are largely related to Basourin. Tragacanthin is considered a neutral polysaccharide component that consists of sequential d-galactose units, and the branched- chains of L-Arabinose are joined to it. There are significant amounts of Methoxyl groups in its structure.
| Texture | Organic material (%) | pH | EC ds.m-1 | Available potassium (ppm) | Available phosphorous (ppm) | Nitrogen total (%) |
|---|---|---|---|---|---|---|
| Loamy Sand | 0.8 | 7.6 | 0.41 | 194.5 | 1.3 | 0.09 |
Table 1: Characteristics of the used soil in experiment.
Study Traits
Measurement of Hydrogen Peroxide Content: The amount of hydrogen peroxide was determined based on the reaction of hydrogen peroxide with potassium iodide (KI) and according to the method of Alexieva V, et al. [18]. In this method, 0.5 g of fresh leaf tissue (the youngest leaves of the stem end) was dissolved in 0.1% Trichloroacetic acid (TCA) at a cold temperature. The extract was centrifuged for 15 minutes at 12,000 rpm. Then, 500 μl of 100 mM potassium phosphate buffer (pH=7) and 2 ml of 1 M potassium iodine were added to 500 μl of the supernatant solution. The reaction mixture was kept at room temperature for 1 hour in the dark, and then the absorbance of the samples was measured at 390 nm. A standard curve was used to calculate the hydrogen peroxide value.
Superoxide Dismutase: In this regard, the method of Dhindsa RH, et al. [19] was used. The basis for measuring this method is SOD enzyme inhibition from the optical reduction of Nitro blue tetrazolium chloride (NBT). The reaction solution consisted of 3 milliliters of 50 mM phosphate buffer (pH = 7.8), 13 mM methionine, 75 microliters of NBT, 2 mM EDTA, and 50 microliters of extracted protein, which were mixed together and in the next step were poured inside the micro- tube. Riboflavin was added at the last stage and the micro- tubes were placed on a shaker and positioned at a distance of 30 cm from a 15 watt fluorescent lamp for 10 minutes. The control sample did not have an enzyme extract. The absorbance was read at 560 nm with a spectrophotometer. Finally, the amount of SOD activity was measured in terms of units per minute per milligram of protein.
Peroxidase: To measure the activity of the peroxidase enzyme, we used the method of Adam MJW, et al. [20]. In this method, the peroxidase enzyme activity was calculated using a 20 mM sodium phosphate buffer (pH = 6) and 200 mM guaiacol as an electron donor. We also used 10 μL of hydrogen peroxide (30% w/v) as an electron receptor. The activity was expressed in unit/mg protein. We read the absorbance using a spectrophotometer at 475 nm for 120 seconds.
Membrane Lipid Peroxidation (Using Malondialdehyde Index): The membrane lipid peroxidation was measured using the method of Davey MW, et al. [21]. A quantity of 0.5 g of fresh leaf samples (the youngest leaves at the stem end) was crushed with liquid nitrogen for 1 min using a pounder.
The crushed leaf powder was then poured into a test tube, and 5 ml of potassium phosphate buffer 50 mM (pH=7) placed in an ice container was added.
The sample was centrifuged at 14000 rpm for 30 minutes at a temperature of 4°C. Then, 1 ml of Thiobarbituric acid 0.5% containing Trichloroacetic acid 20% (w/v) was added to 1 ml of the supernatant solution after centrifugation. The mixture was heated in a hot water bath at 95°C for 30 minutes. To stop the reaction, the container containing the heated mixture was quickly placed in an ice bath for 30 minutes. The cooled mixture was then centrifuged at 10,000 rpm for 10 minutes. The absorbance of the mixture was measured by a spectrophotometer at two wavelengths of 532 and 600 nm.
Soluble Proteins of Leaf: One gram of fresh leaf tissue was sampled to extract the soluble proteins. It was then thoroughly ground with 5 ml of 0.05 molar tris HCL buffer at pH 7.5 at 0-4°C. The resulting solution was centrifuged for 25 minutes at 10,000 rpm at 4°C [22]. The supernatant was used to measure the concentration of soluble protein. In this method, 990 μl of Bradford reagent (including 0.1 g of Coomassie Blue, 50 ml of 96% ethanol, and 100 ml of 85% phosphoric acid) was mixed with 10 μL of Tris buffer to calibrate the spectrophotometer, then calibrated by a spectrophotometer at 595 nm. To measure the amount of soluble proteins, 10 μl of the test sample (extract) was added to 990 μL of Bradford reagent and the absorbance was read at 595 nm. Finally, the amount of soluble proteins was calculated using equation (1) with the aid of absorbance in the standard solution.
( ) ( ) ( ) ( ) Protien mg gfresh weight samplesize ppm calculatedprotiencontent fresh weightof sample 1000 = × × Proline Content: The proline content of the plant was measured using the method described by Bates LS, et al. [23]. First, 0.5 grams of fresh black cumin leaf was crushed with liquid nitrogen in a pounder. The crushed leaf was then transferred to a tube and 10 ml of 3% sulfosalicylic acid was added. The sample was placed on ice and centrifuged at 15,000 rpm for 10 to 15 minutes at 4°C to remove any excess material. Next, 2 ml of the filtered extract was transferred to a microtube. To this, 2 ml of Ninhydrin and 2 ml of glacial acetic acid were added and mixed well. Simultaneously, 2 ml of standard solutions containing 0, 4, 8, 12, 16, and 20 mg/l of proline were also added to separate microtubes. To each microtube, 2 ml of Ninhydrin acid and 2 ml of glacial acetic acid were added and mixed well. The samples were then heated in a hot bath for 1 hour and subsequently placed in an ice bath. To the solution, 4 ml of toluene was added and vortexed for 20 seconds. The proline standards, which were soluble in the toluene phase, were transferred to a cuvette in the spectrophotometer. The proline content was measured at 520 nm and a standard curve was plotted. Finally, the absorption value of the plant samples was determined and inserted into the linear equation (Equation 2) to obtain the proline value.
$$ X = \left[ \left(A. B\right) / C \right] / \left(D / 5\right) $$
Equation 2 In the above equation, X is the proline content of tissue in micrograms/gram of dry tissue, A is the amount of proline obtained from the standard chart in micrograms/ml, B is the amount of toluene used in ml, C is the proline molecular weight, and D is the sample plant weight in grams.
Soluble Carbohydrates (in Water and Alcohol): In order to measure the soluble carbohydrates, first 10 ml of 96% ethanol was added to 0.1 g of leaf and placed in a water bath at 100°C until 2/3 of the alcohol is evaporated. In the next step, alcohol is added again, and then it is centrifuged for 5 minutes at 5300 rpm. After the centrifugation, the solution is separated from the residue (at this stage, the solution is prepared to measure alcohol-soluble carbohydrates). After separating the solution from the residue, the residue is dried in an oven and 10 ml of distilled water is added to it and placed in a water bath at 100°C for 2 hours. In the next step, distilled water is added again, and then it is centrifuged for 5 minutes at 5300 rpm. After the centrifugation, the solution is separated from the residue (at this stage, the solution is prepared to measure water-soluble carbohydrates). The next steps are the same for water and alcohol soluble carbohydrates. By adding 3 ml of Anthrone to 0.5 ml of extract and placing it in a water bath at 100°C for 8 minutes, the process continues. The samples are then placed immediately in ice water and read by a spectrophotometer with a wavelength of 630 nm [24].
Leaf Osmotic Potential: In order to measure osmotic potential, after sampling, the leaves were divided into small pieces and 0.3 grams of each leaf were inserted into a sampler tip pierced with a needle. The sampler head was then placed inside a micro tube (1.5 mL). It was then centrifuged for 20 minutes at 25°C and 13500 rpm. This action allowed the leaf extract to enter the micro tube from the sampler tip. For reading, 1980 μl of distilled water was mixed with 20 μl of the extract and then centrifuged for 10 minutes at 25°C and 14000 rpm. Finally, the osmotic potential of 150 μL of the mixture was read by the osmometer (KNAUER, K-7400, Germany).
Grain Yield: After the black cumin ripened, the grain yield of each pot was harvested separately and the grain yield for each treatment was reported in grams per plant.
Results
Hydrogen Peroxide
Hydrogen peroxide is one of the most important ROSs, and its production is increased under stress conditions, which leads to oxidative damage [25]. As shown in Figure 1, the highest amounts of hydrogen peroxide were produced in all spray treatments in plants under drought stress, and with increasing drought stress, production and accumulation of hydrogen peroxide increased. The lowest levels of hydrogen peroxide production belonged to fully irrigated plants. The results showed that the effect of tragacanth gum on the production of hydrogen peroxide in black cumin was significant.
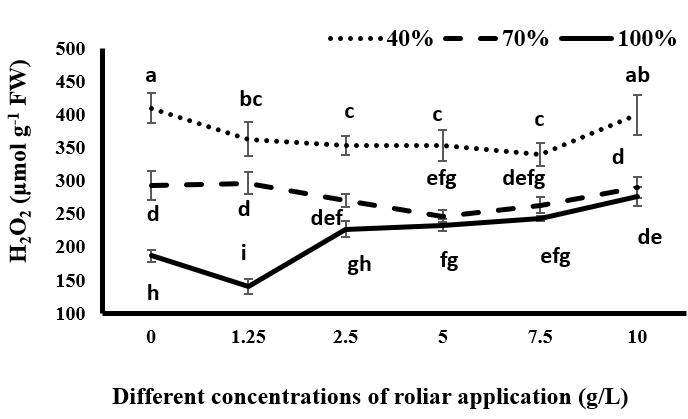
In the full irrigation treatment, the use of tragacanth at a concentration of 1.25 g/L resulted in a decrease in the production of hydrogen peroxide compared to non-sprayed plants. With increasing concentrations of tragacanth material application higher than 1.25 g/L, the production of hydrogen peroxide increased significantly. So, the amount of hydrogen peroxide at 5 g/L and above was significantly increased in comparison to non-consumption of tragacanth. In plants under moderate drought stress, spraying of tragacanth gum up to 5 g/L decreased the production of hydrogen peroxide. But at higher concentrations of tragacanth, the production of hydrogen peroxide increased. Under severe drought stress, application of tragacanth gum up to 7.5 g/L reduced the production of hydrogen peroxide compared to non- application of tragacanth. However, there was no significant difference between the amounts of tragacanth until the mentioned concentration. By increasing the concentration of tragacanth usage higher than 7.5 g/L, the production of hydrogen peroxide increased, but in both moderate and severe drought stress, the amount of hydrogen peroxide production in the consumption of 10 g/L had no significant difference with non-consumption of tragacanth (Figure 1). Therefore, it seems that under drought stress, spraying of black cumin with tragacanth gum is useful in reducing the production of hydrogen peroxide in the plant, especially at lower concentrations. Probably, tragacanth at lower concentrations reduces plant transpiration by reducing water loss from stomata. This increases the water potential of the plant and prevents drought stress occurrence and oxidative stress in the plant. However, complete blockage of stomata at higher concentrations of tragacanth usage, which may occur depending on the amount of available water at different concentrations, reduces the potential and relative water content of the plant, and finally causes the occurrence of drought stress in the plant and increases the production of hydrogen peroxide. Under drought stress conditions, the reduction of CO2 assimilation by closing the stomata results in non-consumption of products obtained from the electron transport chain (NADPH and ATP), and the reduced ferredoxin is increased, which leads to an increase in producing active radicals such as H2O2 [26]. Also, in drought stress conditions due to the limited absorption and stabilization of CO2 and increased activity of the oxygenase of the Rubisco enzyme, photorespiration increases in C3 plants, which will also lead to an increase in H2O2 [27]. Many researchers have linked 70% H2O2 produced during drought stress to photorespiration [28, 29].
Superoxide Dismutase and Peroxidase
In this study, drought stress increased the activity of SOD enzymes Figure 2 and POD Figure 3 in black cumin, which could be due to increased activity of free radicals such as hydrogen peroxide Figure 1 under stress conditions. As shown in Figures 2 & 3 the lowest levels of SOD and POD activity were observed in all treatment groups of tragacanth gum under full irrigation conditions. With the occurrence of drought stress and increasing stress, the activity of these two enzymes significantly increased in all tragacanth usage treatments. In fact, as the stress intensity increased, the production rate of ROS increased and the plant increased its antioxidant enzymes to deal with the produced free radicals. Superoxide dismutase is the first and most important enzyme in the detoxification process of ROS compounds, playing a vital role in cellular defense mechanisms against the risk of hydroxyl radical (OH) formation by converting superoxide radical (O-2) to H2O2 in the cytosol, chloroplasts, and mitochondria [30]. Although hydrogen peroxide is toxic in high concentrations and eliminated from the glutathione- ascorbate cycle by catalase and ascorbate peroxide enzymes, it acts as a messenger in the transmission process and activates genes dependent on the plant’s resistance when present in low concentrations [31]. Chiyaneh RE, et al. [32] stated that the accumulation of SOD and POD is increased in black cumin under drought stress. Under complete irrigation conditions, spraying with 1.25 gr/L significantly reduces the activity of SOD and POD compared to the control group without tragacanth consumption, while the use of higher concentrations of tragacanth significantly increases the activity of the enzymes compared to the control group. The maximum activity of the enzymes was observed at a consumption of 10 g/L tragacanth (Figures 2 & 3). Under mild drought conditions, the use of tragacanth leads to a decrease in the activity of SOD and POD, which was significant at concentrations of 5 and 7.5 g/L tragacanth. The activity of the two enzymes increased with a consumption of 10 g/L tragacanth, placing it in a statistical group with no tragacanth treatment.
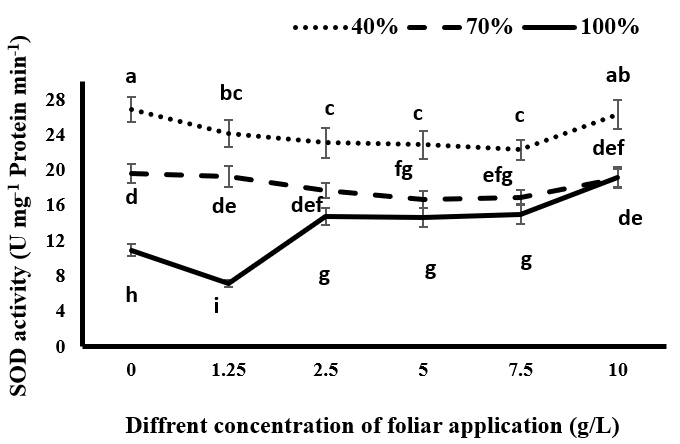
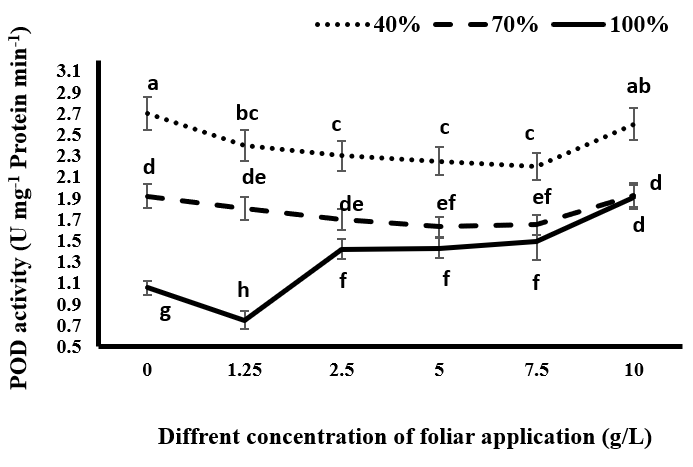
In severe drought stress, the use of different concentrations of tragacanth, except for the concentration of 10 g/L, significantly decreased the activity of SOD and POD. In other words, the use of tragacanth gum in concentrations higher than 7.5 g/L increased the activity of the enzymes compared to lower concentrations, so there was no significant difference between the treatment without tragacanth consumption and the treatment with 10 g/L tragacanth consumption. It seems that at different levels of irrigation, changes in the activity of these enzymes depend on changes in the production of active oxygen species. In the present experiment, the change in the activity of enzymes was consistent with the changes observed in the measured H2O2 (Figure 1).
Malondialdehyde
The minimum amount of MDA was obtained in all concentrations of tragacanth gum application under complete irrigation conditions. Drought stress resulted in an increase in MDA. In all levels of tragacanth consumption, the maximum amount of MDA was obtained under severe drought stress (40% irrigation) (Figure 4). Darvizheh H, et al. [33] also stated that drought stress increased the activity of SOD, POD, and MDA. The plant spraying with tragacanth gum (concentrations higher than 1.25 g/L) led to an increase in MDA compared to the control group (without spraying). However, only 10 g/L of tragacanth significantly increased MDA under complete irrigation conditions. In drought stress conditions (moderate and severe), the consumption of tragacanth reduced MDA amount, but under severe drought conditions and with the use of 7.5 g/L of tragacanth, the reduction was significant compared to non-tragacanth consumption. It can be stated that under drought conditions, spraying black cumin with tragacanth gum (up to 7.5 g/L) led to a decrease in lipid peroxidation in the membrane.
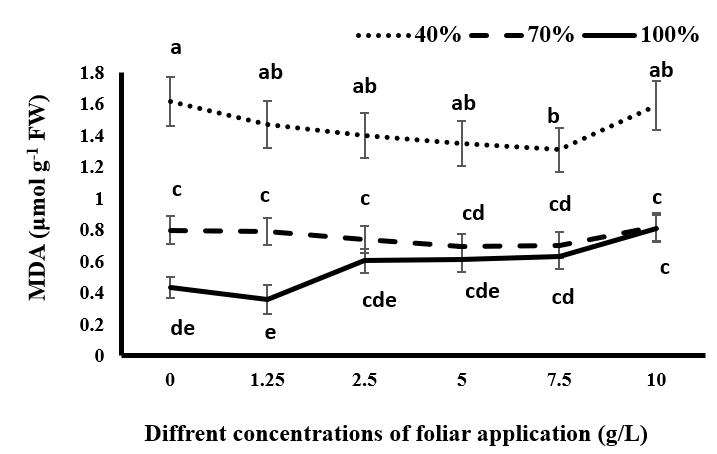
It seems that these concentrations of tragacanth result in a reduction in plant transpiration by decreasing water loss from stomata, which increases the water potential of the plant, reduces the effects of drought stress, and subsequently decreases oxidative stress and lipid peroxidation in the plant. However, complete blockage of stomata at high concentrations of tragacanth, which may occur depending on the available water content at different concentrations, can increase drought stress in the plant and increase the production of hydrogen peroxide Figure 1 and peroxidation of membrane lipids. One of the reactions that is more rapid in the presence of reactive oxygen species is the peroxidation of membrane lipids. In this study, by increasing drought stress, the accumulation of H2O2 is increased (Figure1). Considering the increased reactive oxygen species and their effect on lipid peroxidation (due to the effect on the double bond of unsaturated fatty acid), the degradation of membrane fatty acids is increased [34]. Increasing the activity of SOD Figure 2 and POD Figure 3 enzymes indicates the activation of the antioxidant capacity of the plant against drought-induced oxidative stress (increased H2O2 accumulation). However, in this study, this antioxidant capacity in black cumin was not enough to cope with drought stress because the increase in drought stress resulted in an increase in MDA (Figure 4). Therefore, it can be stated that in many cases, the antioxidant capacity of the plant alone and without managerial approaches will not be able to eliminate all oxidants. In this study, although the use of tragacanth gum did not have a direct effect on the increase of plant antioxidant capacity, it was able to reduce the production of MDA by reducing the production of hydrogen peroxide (Figure 1).
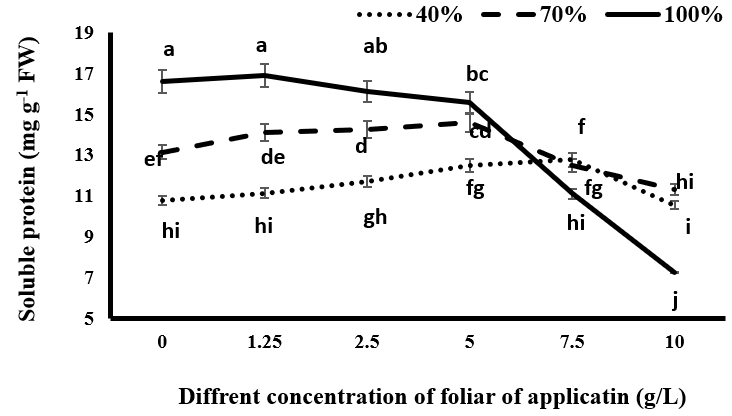
Soluble Proteins
The results showed that the maximum soluble protein content in the leaf was observed under complete irrigation conditions for both the tragacanth application and non- application groups (up to 5 g/L) (Figure 5). Under complete irrigation conditions, increasing the concentration of tragacanth gum led to a decrease in the amount of soluble proteins. Specifically, an increase in tragacanth concentration from 2.5 g/L to higher concentrations significantly decreased this trait, with the lowest amount of leaf soluble proteins (7.26 mg/g of fresh leaf) observed at a tragacanth consumption level of 10 g/L under the same irrigation level. This suggests that the use of 2.5 g/L and higher concentrations of tragacanth resulted in limited stomata turnover and the appearance of drought stress effects on the plant, leading to a decrease in the amount of soluble protein in the leaf. In mild drought stress conditions, the application of tragacanth and an increase in its concentration resulted in an increase in soluble protein content compared to non-tragacanth application. There was a significant difference in concentrations of 2.5 g/L and 5
g/L. However, increasing the tragacanth concentration from 5 g/L to higher concentrations led to a decrease in soluble proteins. At this level of irrigation, the treatment with 10 g/L tragacanth gum had the lowest amount of soluble proteins, which was significantly lower than the soluble protein content in plants without tragacanth consumption. In severe drought stress conditions, consuming tragacanth and increasing its consumption rate up to 7.5 g/L increased the content of soluble proteins. This increase was significant at concentrations of 2.5 g/L, 5 g/L, and 7.5 g/L. However, at a concentration of 10 g/L, the soluble protein content decreased and was statistically similar to the soluble protein content in the treatment without tragacanth application.
Under drought stress, spraying Nigella sativa with tragacanth gum, especially at lower concentrations, improved the production of soluble proteins. Tragacanth gum at relatively low concentrations may reduce water loss through transpiration from stomata, thereby increasing the water potential of the plant and reducing the decomposition of soluble proteins during induced oxidative stress or reducing their production. However, complete blockage of stomata at higher concentrations of tragacanth material, which may occur at different concentrations depending on the water state of the plant, may cause or intensify drought stress in the plant and reduce soluble protein content. Kabiri R, et al. [12] reported a reduction in soluble protein content in black cumin under drought stress. Reduction in soluble protein concentration is a common symptom induced by oxidative stress and is often observed under drought stress. This decrease in protein content under drought stress can be attributed to a lack of protein synthesis or the breakdown of proteins due to increased protease activity [35].
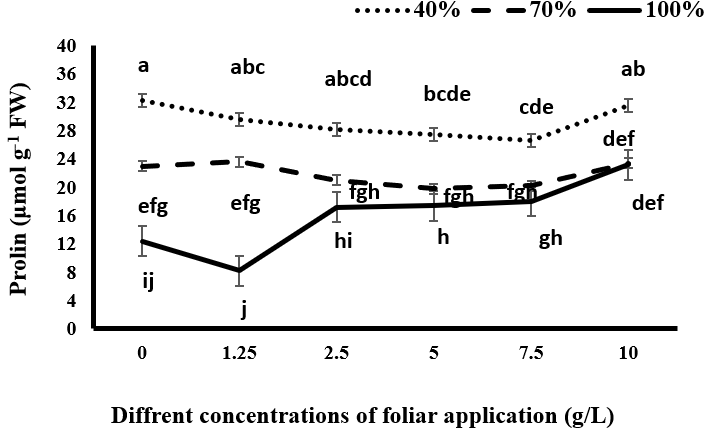
Proline Content
The results indicated that the occurrence of drought stress and increasing its severity leads to an increase in proline content. The maximum amount of proline belonged to plants that were subjected to drought stress, especially severe drought stress. Increasing the amount of proline under drought stress Figure 6 may be related to its osmotic property and its antioxidant property under stress conditions. Proline is accumulated in a wide range of organisms (from bacteria to plants) as it encounters non-biological stresses. Proline synthesis plays a role in decreasing the cytoplasmic osmotic potential and maintaining the NADP+/NADPH ratio. Proline also acts as an osmolyte, remover of free radicals, stabilizer of macromolecules, and a cell wall component. The importance of proline accumulation in preserving the plant’s water status is more important than other organic materials, and proline acts as the most common accumulated osmolyte under stress conditions [13]. Proline plays a key role in the stability of proteins and cell membranes during osmotic stresses [36]. Proline, as a chemical protector, stabilizes the natural form of proteins, prevents the deformation of the natural form of enzymatic compounds, chelates the metals, prevents the peroxidation of lipids, and preserves the integrity of the membrane. Therefore, during drought stress, proline production is increased to protect the plant against oxidative stress [13]. Darvizheh H, et al. [33] stated that the proline content in the plant is increased during drought stress. In mature leaves, protein decomposition leads to a reduction in their concentration and, as a result, an increase in the free amino acids, including proline [35].
Spraying black cumin with different concentrations of tragacanth gum under various irrigation levels had varying effects on the proline content of the plant. Under complete irrigation conditions, the use of 1.25 g/L tragacanth resulted in a reduction in proline content, although this reduction was not significant. Increasing the concentration of tragacanth application led to an increase in proline content, with significant increases observed at concentrations of 5, 7.5, and 10 g/L. The highest proline content was observed at the 10 g/L concentration of tragacanth gum. Under mild drought conditions, spraying the plant with tragacanth gum had no significant effect on proline content. However, under severe drought stress, the consumption of tragacanth gum up to 7.5 g/L resulted in a reduction in proline content compared to non-sprayed plants. This reduction was significant only at concentrations of 5 and 7.5 g/L of tragacanth gum. At this irrigation level, the use of 10 g/L tragacanth gum increased proline content compared to other concentrations, placing it in the same statistical group as non-sprayed plants. Under complete irrigation conditions, where the plant does not face water limitations, the use of tragacanth gum, especially at concentrations of 1.25 g/L or higher, led to the observation of drought-like effects in the plant. This is due to the blocking of stomata, which prevents transpiration and reduces the plant’s ability to absorb water. As a result, proline content increases through synthesis or protein decomposition.
In drought stress conditions, spraying black cumin with up to 10 g/L tragacanth gum was found to be useful in reducing plant transpiration and improving the water status of the plant, as well as reducing the amount of proline. However, using a concentration of 10 g/L tragacanth increased stomatal constraints and diminished the beneficial effects of tragacanth on drought stress adjustment in the plant. Stomata play a crucial role in the photosynthesis process by facilitating both water exit and carbon dioxide absorption. The initial absorption of carbon dioxide occurs through the stomatal openings alongside water outflow. Therefore, when water flow is limited (such as with high levels of anti- transpiration compounds), the amount of carbon dioxide absorption decreases [37]. The use of anti-transpiration compounds under optimal irrigation conditions can hinder the growth and photosynthesis process of the plant due to their adverse effects on gas exchange [38].
Soluble Carbohydrates in Water and Alcohol
As seen in Figures 7 & 8, drought stress has increased the accumulation of soluble carbohydrates in water and alcohol in the plant. Increasing the intensity of drought stress has also increased the accumulation of these compounds in the plant. Plants produce low molecular weight soluble materials such as amino acids, sugars, and organic acids to regulate intracellular osmotic ability under adverse environmental conditions such as drought stress. Proline and soluble carbohydrates are the most common compounds and play a role in maintaining cellular swelling and adaptation under drought stress conditions. Proline and soluble carbohydrates also help remove free oxygen radicals from plant cells and reduce the effects of drought stress on physiological responses such as the opening and closure of stomata and photosynthesis by increasing the osmotic concentration [39].
Under complete irrigation conditions, although the consumption of 1.25 g/L tragacanth gum reduced the content of alcohol-soluble carbohydrates, this was not significant. By increasing the concentration of tragacanth gum, the content of soluble carbohydrates in water and alcohol increased. This increase was significant at concentrations of 7.5 and 10 g/L tragacanth for alcohol-soluble carbohydrates and 10 g/L for water-soluble carbohydrates compared to the control without spraying. At this level of irrigation, the maximum content of soluble carbohydrates was observed at a 10 g/L tragacanth concentration. Under mild stress conditions, the spraying of black cumin with tragacanth gum had no significant effect on the content of carbohydrates soluble in water and alcohol. Under severe drought stress, tragacanth application and increasing its concentration up to 7.5 g/L reduced the content of soluble carbohydrates in the plant compared to control plants. However, this decrease was only significant at the concentration of 7.5 g/L for carbohydrates soluble in water and at the concentrations of 5 and 7.5 g/L for carbohydrates soluble in alcohol. At this level of irrigation, the consumption of 10 g/L tragacanth compared to other concentrations increased the water and alcohol soluble carbohydrates in black cumin. As a result, they were placed in a statistical group with non-sprayed plants in terms of the considered traits.
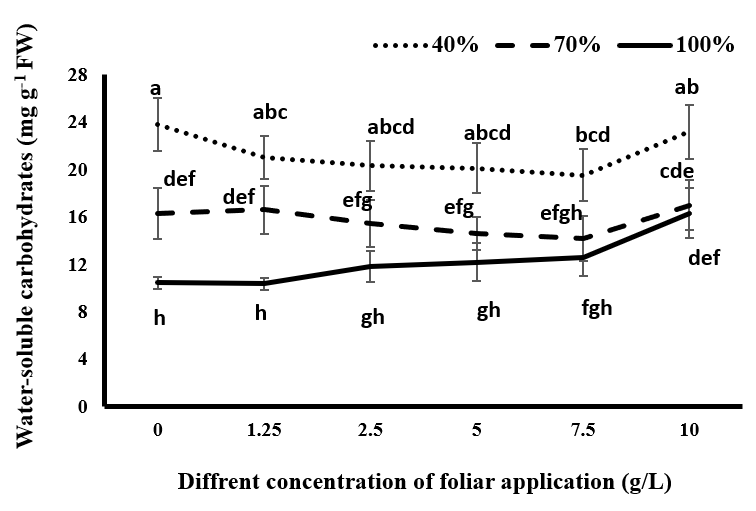
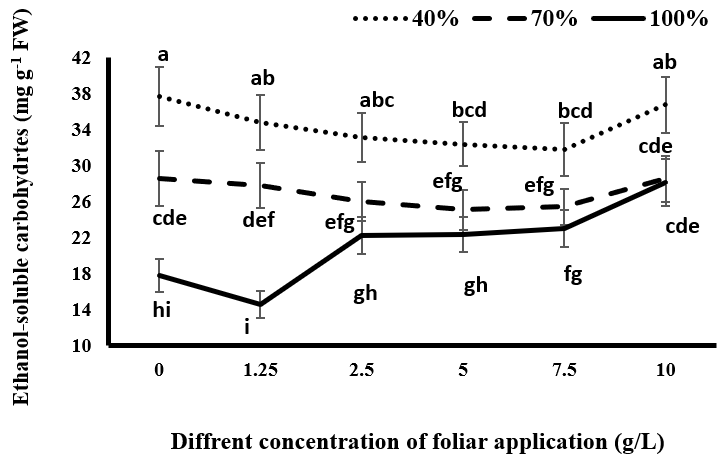
Under complete irrigation conditions, the use of tragacanth, especially at concentrations higher than 1.25 g/L, probably reduces the opening of stomata and reduces the amount of water absorption required by the plant. This observation suggests that drought stress effects in the plant are observed. As a response to drought stress, the plant increases the amount of soluble carbohydrates to cope with it and increases the osmotic potential to maintain the water potential inside the plant. In drought stress conditions, spraying the plant with up to 10 g/L tragacanth gum decreases the production of soluble carbohydrates due to a reduction in plant transpiration and improvement in water conditions within the plant. However, the use of 10 g/L tragacanth reduces the beneficial effect of tragacanth material on moderating drought stress in the plant by increasing stomata limitation. Due to increased drought stress at this concentration, the accumulation of soluble carbohydrates in the plant is increased compared to other concentrations of tragacanth application.
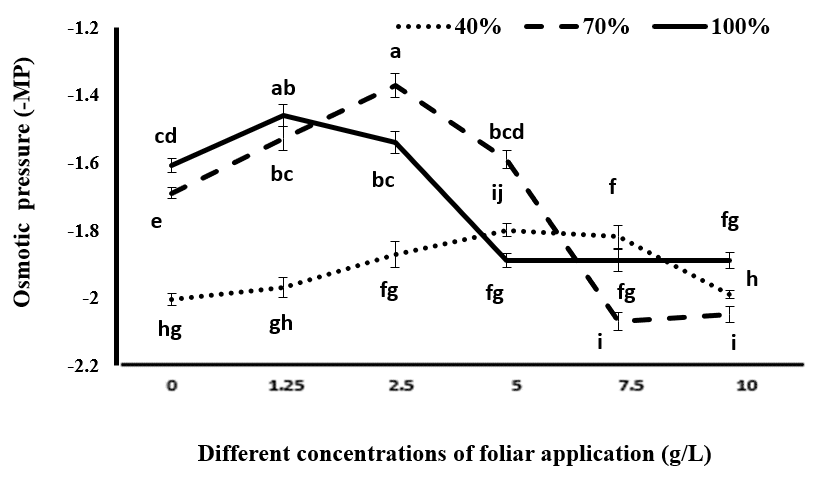
Osmotic Potential
As shown in Figure 9, the osmotic potential of the plant increased with increasing drought stress. The lowest osmotic potential was obtained with irrigation at 70% of the field capacity and a use of 2.5 g/L of tragacanth gum, as well as with full irrigation and a consumption of 1.25 g/L of tragacanth gum. The effect of different concentrations of tragacanth material on the osmotic potential of the leaf varied under different irrigation regimes. Under full irrigation, consumption of tragacanth gum up to 1.25 g/L reduced osmotic potential, but concentrations higher than 1.25 g/L increased the more negative osmotic potential of the plant. Under irrigation at 70% of the field capacity, consumption of tragacanth gum up to 2.5 g/L reduced the osmotic potential, and concentrations higher than 2.5 g/L resulted in a more negative osmotic potential of the plant. Under irrigation at 40% of the field capacity of soil, increasing the concentration of tragacanth gum up to 5 g/L reduced osmotic potential, which was not significantly different from treating the plant with a concentration of 7.5 g/L of tragacanth gum. However, at a higher concentration (10 g/L), the osmotic potential increased, becoming more negative (Figure 9). These changes in osmotic potential indicate the positive effects of optimal concentrations of tragacanth as an anti-transpiration agent in reducing the effects of drought stress, and the negative effects of using high concentrations of tragacanth in exacerbating the effects of drought stress on plants. During water shortages, preserving the water potential of the plant is essential for growth, which can be achieved through the mechanisms of osmotic adjustment resulting from the accumulation of compatible solutions such as proline Figure 6 and soluble carbohydrates in water and alcohol Figures 7 & 8 in the cytoplasm [40].
Grain Yield
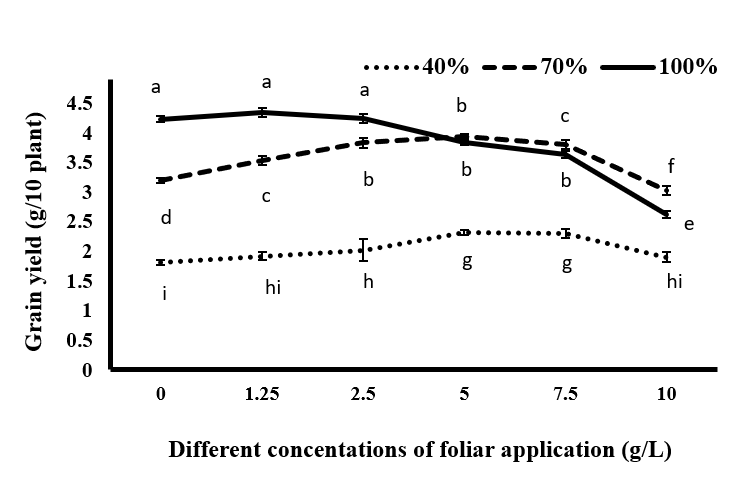
Results showed that the highest grain yield was 4.34, 4.24, and 4.23 g/10 plants, obtained at concentrations of 1.25, 0, and 2.5 g/L of tragacanth gum, respectively, under full irrigation conditions. Drought stress caused a significant decrease in grain yield, with the lowest yield (1.81 g/10 plants) observed in plants under severe drought stress and no tragacanth consumption (Figure 10). In the present research, drought stress resulted in oxidative stress in the plant due to reduced irrigation. This led to an increase in the production of reactive oxygen species, such as H2O2, which in turn increased the peroxidation of membrane lipids and MDA levels in the plant. Vurukonda SS, et al. [41] stated that drought stress leads to the production of active oxygen species, which ultimately reduces grain yield [42].
Increasing activity of SOD (Figure 2) and POD (Figure 3) enzymes indicates the activation of antioxidant capacity of the plant against drought-induced oxidative stress, but this antioxidant capacity in black cumin was not sufficient to cope with drought stress, which increased the accumulation of MDA (Figure 4). Moreover, the plant also increased the amount of proline (Figure 5) and soluble carbohydrates in water (Figure 6) and alcohol (Figure 7) to cope with drought stress, which ultimately led to a negative osmotic potential (Figure 8). In fact, although the accumulation of consistent osmolytes increases the tolerance to drought, this is costly for the plant and indicates the negative effect of stress on the plant’s normal activities. In general, these effects of drought stress reduced photosynthesis, which appeared as grain yield reduction (Figure 9). Therefore, any water shortage stress during growth and development stages of the plant can have a negative effect on the source and reservoir relationships and cause a significant decrease in grain yield [43]. Reduced yield of Nigella sativa during drought stress has also been reported by other researchers [7, 32].
Also, the comparison of mean values of grain yield showed (Figure 9) that spraying of black cumin with tragacanth material significantly increased the grain yield under drought stress conditions. Under the complete irrigation treatment, consumption of 1.25 g/L of tragacanth increased the grain yield, although this increase was not significant, and the grain yield obtained for the sprayed plant by 1.25 and 2.5 g/L of tragacanth showed no significant difference with the non-sprayed plant. In this level of irrigation, increasing the concentration of tragacanth from 2.5 g/L to higher concentrations resulted in a significant decrease in grain yield compared to non-sprayed plants, so that at concentrations of 7.5 and 10 g/L tragacanth gum, the grain yield showed significant reduction compared to the other plants subjected to mild drought under the same concentrations of tragacanth. Under mild stress conditions, spraying significantly increased grain yield, so that at this level of irrigation, the highest grain yield was 3.94 g in 10 plants at 5 g/L of tragacanth. The plant spraying with this concentration of tragacanth increased the grain yield to 23.1% compared to control without tragacanth, and even the amount of the obtained product was significantly higher than that of black cumin at the same concentration of tragacanth consumption under full irrigation conditions (84.3 g in 10 plants). Under severe drought stress, the tragacanth consumption up to 7.5 g/L increased grain yield and application of 10 g/L of tragacanth resulted in a decrease in grain yield, although there was no significant difference with non-application of tragacanth. In order to justify the positive effects of tragacanth gum spraying on grain yield in different irrigation regimes, it can be stated that grain yield was significantly affected by the effect of different concentrations of tragacanth consumption on the amount of production of hydrogen peroxide (Figure 1), the amount of MDA (Figure 4), activity of superoxide dismutase enzymes (Figure 2) and peroxidase (Figure 3), the accumulation of compatible osmolytes (Figures 5-7) and osmotic potential (Figure 8). Application of suitable concentrations of tragacanth gum increased the grain yield by increasing the plant’s osmotic potential and improving the water status of the plant and reducing the production of hydrogen peroxide and malondialdehyde. The studies performed on the use of anti-transpiration agents stated the reducing effects of drought stress and preventing the reduction of yield due to the use of anti-transpiration agents [44, 45, 46, 47].
Correlation among Traits
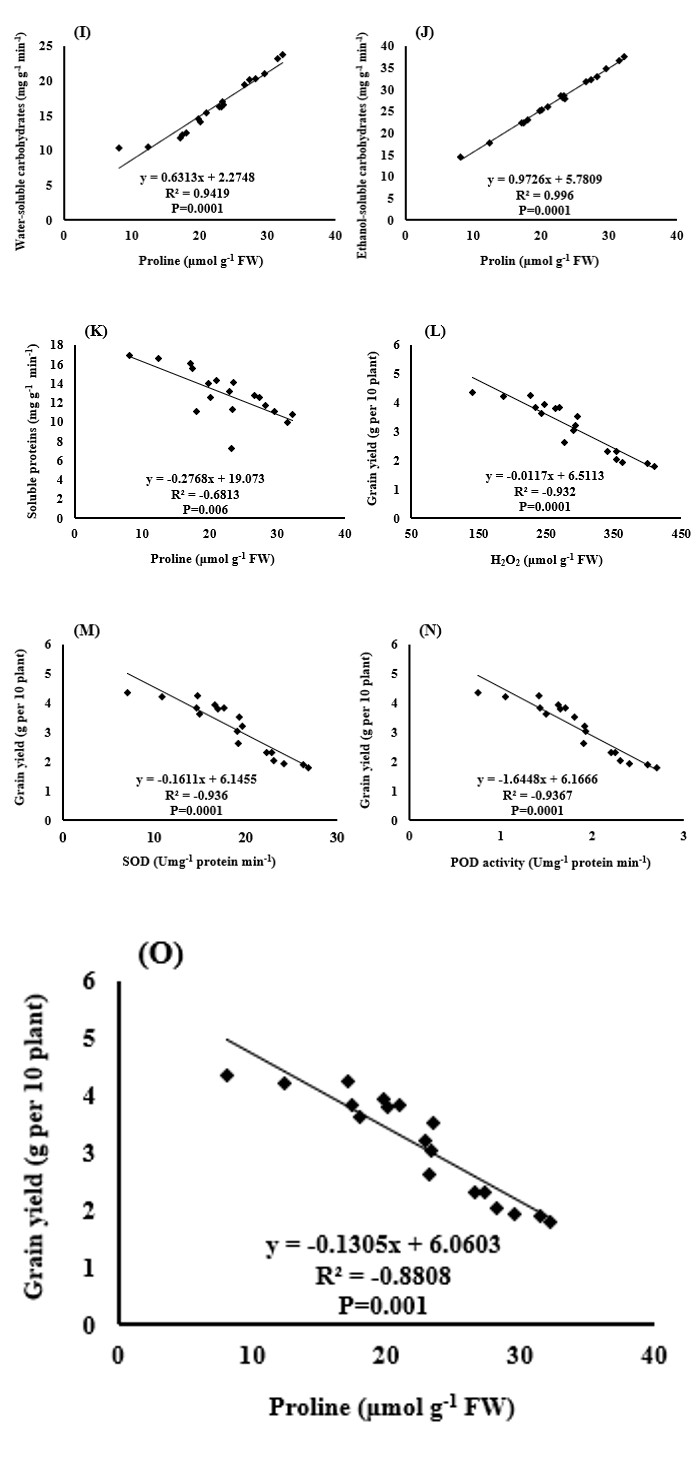
The results showed that the correlation between the amount of hydrogen peroxide produced in the plant and the amounts of malondialdehyde, SOD and POD activity, proline content, and soluble carbohydrates in water and ethanol were completely positive and significant (Figures 11A-11F). But there was a negative correlation between hydrogen peroxide and soluble proteins of the leaf (Figure 11G). These results indicate that increasing the production and accumulation of reactive oxygen species, such as H2O2, increases the membrane lipid peroxidation and consequently increases MDA, as well as the degradation of soluble proteins. The plant increases antioxidant enzymes (SOD and POD) activity and compatible osmolytes (proline and carbohydrates soluble in water and ethanol) to cope with oxidative stress, thereby reducing the destructive effects of reactive oxygen species. There was also a positive and significant correlation between the amount of MDA and the activity of the SOD and POD enzymes and proline content of the plant (Figures 11H-11IJ, which in fact confirms that by increasing the hydrogen peroxide and destructive effects of this oxidant and other reactive oxygen species on the cell membrane, the plant resists against oxidative stress by increasing the activity of antioxidant enzymes and the production of compatible and protective osmolytes [48, 49, 50].
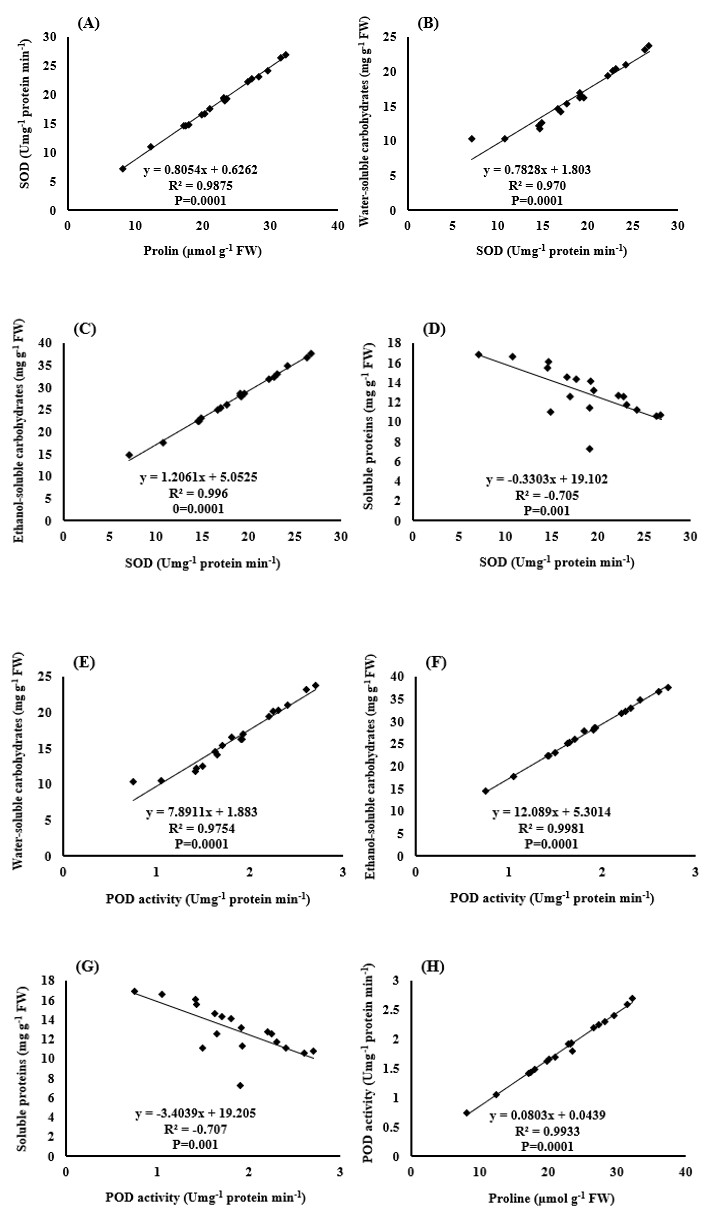
The results also showed that there was a positive and significant correlation between enzyme activity and proline content and soluble carbohydrates in water and ethanol (Figures 12A-12H). In fact, these enzymes and plant compounds (proline and soluble carbohydrates) form part of the protective mechanisms of the plant against drought stress and oxidative stress caused by it. So it is natural to see a positive correlation between them (Figures 12I,12J).
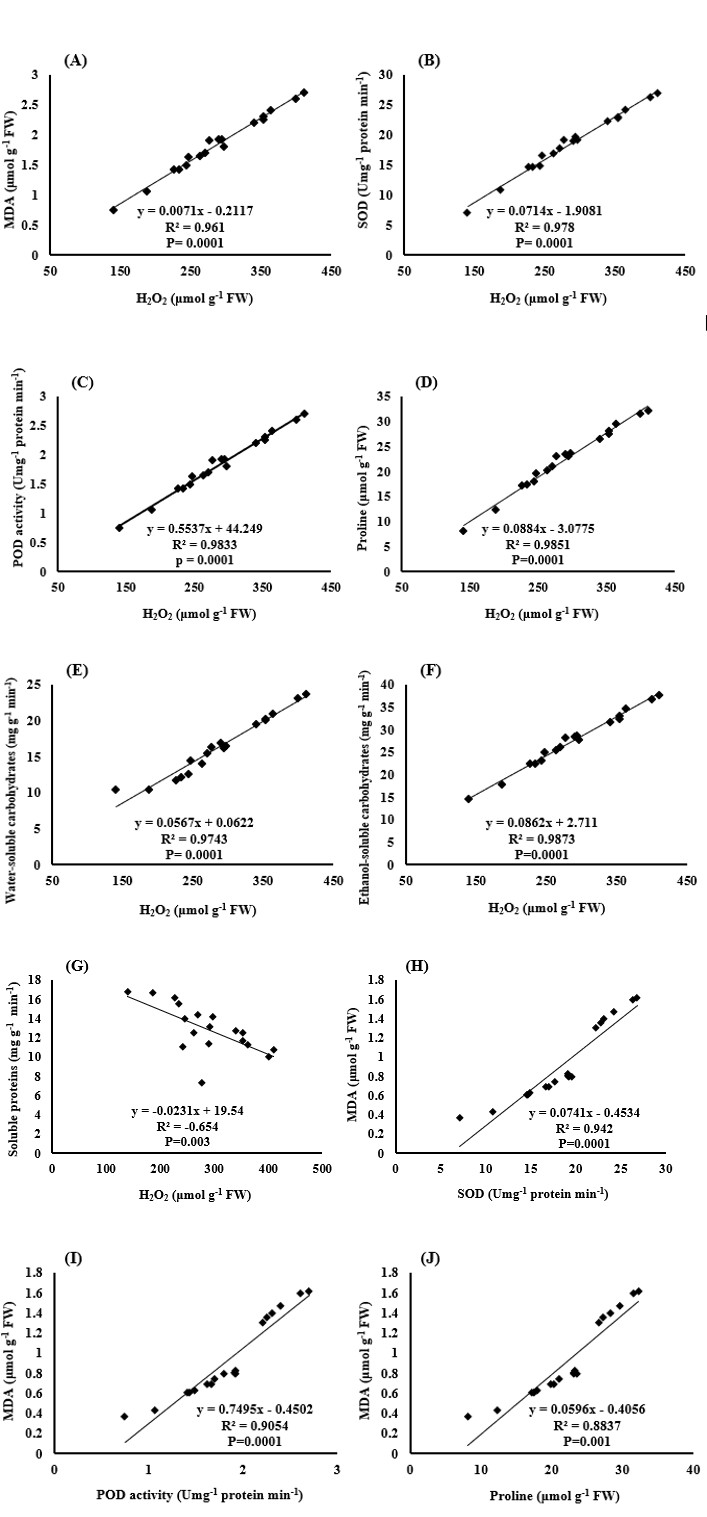
The correlation between the amount of soluble proteins in the leaves and other measured traits, such as the activity levels of the SOD and POD enzymes and the proline content of the plant (Figures 12D,12G & 12K), was negative. In interpreting this negative correlation, it can be stated that the degradation of soluble proteins is part of the oxidative stress damage that results in the plant’s reaction by increasing the activity of antioxidant enzymes and increasing the production of osmolytes, such as proline [51, 52].
Finally, there was a negative and significant correlation between the amount of H2O2, the activity of SOD and POD enzymes, and the proline content of the plant with the grain yield of black cumin (Figures 12L-12O). This negative correlation shows that the production and accumulation of reactive oxygen species, such as H2O2, leads to the destruction of membrane lipids, photosynthetic pigments (data not shown), and macromolecules, such as proteins. As a result, there is disruption in photosynthesis and respiration activities. On the other hand, the plant copes with induced stress by activating its protective mechanisms and tolerating stress through the production of antioxidant enzymes and osmotic regulators. The production of these enzymes and osmotic compounds in plants requires cost and energy consumption, which causes part of the energy production in the plant to be used instead of the production of grain to deal with stress. Naturally, the result of these metabolic changes in the plant will be a reduction in yield [53].
Conclusion
Under drought stress conditions, the increased production of reactive oxygen species, including H2O2 as a result of oxidative stress, leads to an increase in the peroxidation of lipid membranes. In fact, the protective mechanisms of the plant, such as increasing the activity of antioxidant enzymes SOD and POD, and the production and accumulation of proline and soluble carbohydrates in response to oxidative stress, are not enough to produce, remove, or disable active oxygen species and their destructive effects on macromolecules such as proteins. In such a situation, managerial solutions can change the equation for the benefit of the plant. In the present study, the use of tragacanth gum was able to reduce the effects of drought stress by reducing water losses, improving the water status of the plant, and reducing the effects of oxidative stress. This caused the plant to spend less energy coping with stress, ultimately leading to an increase in grain yield of black cumin. The positive effects of tragacanth consumption on reducing and modifying the effects of drought stress at different levels of irrigation and different concentrations of tragacanth were different. In the present study, under full irrigation conditions, lower concentrations of tragacanth were useful, while in drought stress conditions, higher concentrations of tragacanth (except 10 g/L) were useful. Therefore, it can be stated that tragacanth consumption in optimal concentrations as an anti- transpiration agent can possibly decrease the effects of drought stress in arid, semi-arid, and Mediterranean regions facing water shortage. The effect of tragacanth gum on reducing and modifying the effects of drought stress in different plants requires further studies and extensive research.
Future Research
The application of tragacanth in field conditions and in different plants can be done by researchers. Depending on the severity of the stress, various concentrations of anti- transpiration agents are optimized, and their applications on other crops can also be an option for future research.
References
-
Fallaha S, Malekzadeha S, Pessarakli M (2018) Seed priming improves seedling emergence and reduces oxidative stress in Nigella sativa under soil moisture stress. Journal of Plant Nutrition 41(1): 29-40.
-
Hoerling M, Eischeid J, Perlwitz J, Quan X, Zhang T, et al. (2012) On the increased frequency of Mediterranean drought, Journal of Climate 25: 2146-2161.
-
Hertig E, Tramblay Y (2017) Regional downscaling of Mediterranean droughts under past and future climatic conditions, Global and Planetary Change 151: 36-48.
-
Naumann G, Alfieri L, Wyser K, Mentaschi L, Betts RA, et al. (2018) Global changes in drought conditions under different levels of warming. Geophysical Research Letters 45(7): 3285-3296.
-
Manavalan P, Nguyen HT (2017) Drought Tolerance in Crops: Physiology to Genomics. Plant Stress Physiology 2.
-
Pascoa P, Gouveia CM, Russo A, Trigo RM (2017) The role of drought on wheat yield interannual variability in the Iberian Peninsula from 1929 to 2012, International Journal of Biometeorology 61(3): 439-451.
-
Hadi HSMR, Darzi MT, Riazi GH (2016) Black cumin (_Nigella sativa_ L.) yield affected by irrigation and plant growth promoting bacteria. Journal of Medicinal Plants and By-products 5(2): 125-133.
-
Lotfi R, Pessarakli M, Kouchebagh GP, Khoshvaghti H (2015) Physiological responses of Brassica napus to fulvic acid under water stress: chlorophyll a fluorescence and antioxidant enzyme activity. Crop Journal 3: 434- 439.
-
Shulaev V, Oliver DJ (2006) Metabolic and proteomic markers for oxidative stress, new tools for reactive oxygen species research. Plant Physiology 141(2): 367- 372.
-
Zhang F, Shi W, Jim Z, Shen Z (2003) Response of antioxidative enzymes in cucumber chloroplasts to cadmium toxicity. Journal of Plant Nutrition 26: 1779- 1788.
-
Hasanuzzaman M, Alam MM, Rahman A, Hasanuzzaman M, Nahar K, et al. (2014) Exogenous proline and glycine betaine mediated up regulation of antioxidant defense and glyoxalase systems provides better protection against salt-induced oxidative stress in two rice (_Oryza_ _sativa_ L.) varieties. BioMed Research International. 2014: 757219.
-
Kabiri R, Nasibi F, Farahbakhsh H (2014) Effect of exogenous salicylic acid on some physiological parameters and alleviation of drought stress in _Nigella_ _sativa_ plant under hydroponic culture. Plant Protection Science 50(1): 43-51.
-
Hayat S, Hayat Q, Alyemeni MN, Wani AS, Pichtel J, et al. (2012) Role of proline under changing environments. Plant Signaling and Behavior 7(11): 1456-1466.
-
Chaitanya KV, Rasineni GK, Reddy AR (2009) Biochemical responses to drought stress in mulberry (_Morus alba_ L.): evaluation of proline, glycine betaine and abscisic acid accumulation in five cultivars. Acta Physiology Plantarum 31: 437-447.
-
Xue G, Lynne McIntyre C, Glassop D, Shorter R (2008) Use of expression analysis to dissect alterations in carbohydrate metabolism in wheat leaves during drought stress. Plant Molecular Biology 67: 197-214.
-
Hajihashemi Sh, Sofo A (2018) The effect of polyethylene glycol-induced drought stress on photosynthesis, carbohydrates and cell membrane in _Stevia_ _rebaudiana_ grown in greenhouse. Acta Physiologiae Plantarum 40: 142.
-
Saucedo CCM, Alvarado DD, Tellez CL, Hernandez GV, Varela SA, et al. (2012) Changes in carbohydrate concentration in leaves, pods and seeds of dry bean plants under drought stress. Interciencia journal 37(3): 168-175.
-
Alexieva V, Sergei I, Mapelli S, Karanov E (2001) The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environment 24: 1337-1344.
-
Dhindsa RH, Plumb Dhindsa R, Thorpe TA (1981) Leaf senescence correlated with increased level of membrane permeability, lipid peroxidation and decreased level of SOD and CAT. Journal of Experimental Botany 32: 93- 101.
-
Adam MJW, Nelson CJ, Sharp RE (1992) Peroxidase activity in the leaf elongation zone of tall fescue. Plant Physiology 99(3): 872-878.
-
Davey MW, Stals E, Panis B, Keulemans J, Swennen RL (2005) High throughput of malondialdehyde in plant tissues. Analytical Biochemistry 347(2): 201-207.
-
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72: 248-254.
-
Bates LS, Waldern RP, Tear ID (1973) Rapid determination of free proline for water stress studies. Plant and Soil 39: 205-207.
-
Hedge JE, Hofreiter BT (1962) Carbohydrate chemistry 17. In: Whistler RL, Be Miller JN (Eds.), Academic Press, New York.
-
Wang H, Shabala L, Zhou M, Shabala S (2019) Developing a high-throughput phenotyping. method for oxidative stress tolerance in barley roots. Plant Methods 15: 12.
-
Sun Y, Geng Q, Du Y, Yang X, Zhai H (2017) Induction of cyclic electron flow around photosystem I during heat stress in grape leaves. Plant Science 256: 65-71.
-
Miller G, Suzuki N, Yilmaz CS, Mittler R (2010) Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell and Environment 33(4): 453-467.
-
De Carvalho CM (2008) Drought stress and reactive oxygen species. Plant Signaling and Behavior 3(3): 156- 165.
-
Noctor G, Jovanovic VSONJA, Driscoll S, Novitskaya L, Foyer CH (2002) Drought and oxidative load in the leaves of C3 plants: a predominant role for photorespiration. Annals of Botany 89(7): 841-850.
-
Khatun S, Ali BM, Hahn EJ, Paek KY (2008) Copper toxicity in _Withania somnifera_: Growth and antioxidant enzymes responses of in vitro grown plants. Environmental and Experimental Botany 64(3): 279-285.
-
Unyayar S, Kele Y, Cekic FO (2005) The antioxidative response of two tomato species with different drought tolerances as a result of drought and cadmium stress combinations. Plant Soil Environment 51(2): 57-64.
-
Chiyaneh RE, Seyyedi S, Ebrahimian M, Siavash E, Moghaddam S, et al. (2018) Exogenous application of gamma-aminobutyric acid (GABA) alleviates the effect of water deficit stress in black cumin (_Nigella sativa_ L.) Esmaeil. Industrial Crops and Products 112: 741-748.
-
Darvizheh H, Zahedi M, Abbaszadeh B, Razmjoo J (2019) Changes in some antioxidant enzymes and physiological indices of purple coneflower (_Echinacea purpurea_ L.) in response to water deficit and foliar application of salicylic acid and spermine under field condition. Scientia Horticulturae 247: 390-399.
-
Shahid M, Pourrut B, Dumat C, Nadeem M, Aslam M, et al. (2014) Heavy-metal induced reactive oxygen species: phytotoxicity and physicochemical changes in plants. Reviews of Environmental Contamination and Toxicology 232: 1-44.
-
Bajji M, Lutts S, Kinet JM (2001) Water deficit effects on solute contribution to osmotic adjustment as a function of leaf ageing in three durum wheat (_Triticum durum_ Desf.) cultivars performing differently in arid conditions. Journal of Plant Science 160(4): 669-681.
-
Zaki RN, Radwan TE (2011) Improving wheat grain yield and its quality under salinity conditions at a newly reclaimed soil by using different organic sources as soil or foliar applications. Journal of Applied Science Research 7: 42-55.
-
Davenport DC (1967) Effects of chemical antitranspirants on transpiration and growth of grass. Journal of Experimental Botany 18(55): 332-347.
-
Davenport DC, Hagan RM, Martin PE (1969) Antitranspirant Uses and effects on plant life. California Agriculture 23(5): 14-16.
-
Tiryaki I (2016) Drought stress and tolerance mechanisms in alfalfa (_Medicago sativa_ L.). KSU Journal of Natural Science 19(3): 296-305.
-
Ajithkumar P, Panneerselvam R (2013) Osmolyte accumulation, photosynthetic pigment and growth of _Setaria italica_ (L.) P. Beauv. under droght stress. Asian Pacific Journal 2(3): 220-224.
-
Vurukonda SS, Vardharajula S, Shrivastava M, Skz A (2016) Enhancement of drought stress tolerance in crops by plant growth promoting rhizobacteria. Microbiological Research 184: 13-24.
-
Wei L, Wang L, Yang Y, Wang P, Guo T, et al. (2015) Abscisic acid enhances tolerance of wheat seedlings to drought and regulates transcript levels of genes encoding ascorbate-glutathione biosynthesis. Frontiers in Plant Scince 30: 458-469.
-
Pessarkli M (1999) Handbook of plant and crop stress. USA, pp: 697.
-
Weerasinghe MM, Kettlewell PS, Grove IG, Hare MC (2016) Evidence for improved pollen viability as the mechanism for film antitranspirant mitigation of drought damage to wheat yield. Crop and Pasture Science 67(2): 137-146.
-
Faralli M, Grove I, Hare M, Boyle R, Williams K, et al. (2016) Canopy application of film antitranspirants over the reproductive phase enhances yield and yield- related physiological traits of water stressed oilseed rape (_Brassica napus_ L.). Crop and Pasture Science 67(7): 751-765.
-
Faralli M, Grove IG, Hare MC, Alcalde‐Barrios A, Williams KS, et al. (2017) Modulation of _Brassica napus_ source– sink physiology through film antitranspirant induced drought tolerance amelioration that is dependent on the stress magnitude. Journal of Agronomy and Crop Science 203(5): 360-372.
-
Yan W, Guo S, Li P, Song R, Yu J (2018) Foliar antitranspirant and soil superabsorbent hydrogel affect photosynthetic gas exchange and water use efficiency of maize grown under low rainfall conditions. Journal of the Science of Food and Agriculture 99(1): 350-359.
-
Brillante L, Belfiore N, Gaiotti F, Lovat L, Sansone L, et al. (2016) Comparing Kaolin and Pinolene to Improve Sustainable Grapevine Production during Drought. PLoS ONE 11(6): e0156631.
-
Fallaha M, Eischeid J, Perlwitz J, Quan X, Zhang T, et al. (2012) On the increased frequency of Mediterranean drought. Journal of Climate 25: 2146-2161.
-
Laspina NV, Groppa MD, Tomaro ML, Benavides MP (2005) Nitric oxide protects sunflower leaves against Cd-induced oxidative stress. Plant Science 169(2): 323- 330.
-
Matysik J, Alia A, Bhalu B, Mohanty P (2002) Molecular mechanisms of quenching of reactive oxygen species by proline under stress in plants. Current Science 82(5): 525-532.
-
SAS (2011) SAS for Windows Version 9.3. SAS Institute Inc. Cary NC, USA.
-
Schlegel HG (1956) The utilization of organic acids by chlorella in lincht. Planta 47: 510-515.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note