Transfer of Aminolevulinic Acid-Induced Protoporphyrin IX from Tumor Cells into the Blood
This work evaluates the aminolevulic acid (ALA)-induced Protoporphyrin IX (PpIX) synthesis and the PPIX transference from the tumor cells to the whole blood cells. Human prostate tumor cells (DU145 cells) received ALA at different periods and doses to analyze the PPIX synthesis. Thereafter, blood of healthy animals was added to the cultured tumorigenic cells to monitor the transference of the PPIX synthesized from them to the blood cells. Our results demonstrated that the intracellular production and accumulation of PPIX into prostate human tumor cells depends on the ALA doses and the time of exposition. Besides, PPIX molecules synthesized by tumor cells were transferred to the blood cells, but they were not transferred to the DMEM medium solution, showing that it is not a simple diffusion process of PPIX migration, but probably a complex efflux mechanism that cells have developed to transport it through the blood cells to regulate PPIX concentration inside them, since an excess of PPIX is highly toxic. These findings can be useful to understand the accumulation of PPIX to the blood of tumor-bearing animals and cancer patients, and it can be useful to design treatment and diagnosis protocols.
Introduction
Delta-aminolevulinic acid, ALA, is one of the most selective photosensitizer precursor currently used to diagnosis and treatment of cancer [1, 2, 3, 4]. ALA is an early precursor in heme biosynthesis pathway where it is metabolized to an active sensitizer, the Protoporphyrin IX (PPIX). PPIX has been found to be preferentially accumulated in tumor when compared with normal cells. This phenomenon can be explained by the abnormal metabolism of PPIX in malignant cells. The selective synthesis of PPIX in cancer subjects provide a strong color contrast between the intense red fluorescence of malignant tissue and the weak fluorescence of normal tissue [5, 6, 7]. Abnormal metabolism of PPIX has also been observed in the blood, plasma and erythroid cells of cancerous patients [8, 9, 10]. The mechanism of preferential accumulation of PPIX in the blood of tumor-bearing animals and cancer patient’s remains obscure [11]. Masilamani, et al. [10] proposed that the accumulation of PPIX in the blood occurs when it passes through the tumor, which has lower pH values than normal tissues, and it carries the protonated PPIX. Although the movement of heme/porphyrin into cells has been supposed to occur by a diffusion process, recent studies have described that these compounds are incapable of readily traversing the biological membranes [11], because the negative charges introduced by anionic carboxylate side chains which limit their diffuse across membranes. Some efflux systems that export heme/porphyrin from cells to protect them from the accumulation of excess heme/porphyrin have been proposed [11, 12]. These unsolved questions stimulate our interest to study the transference of ALA-induced PpIX, produced by DU145 cancer cells, from intracellular space to the blood.
Materials and Methods
Cell Line and Cell Culture Conditions
Human prostate tumor cells (DU145 cells) were cultured in DMEM containing high Glucose (4.5 g/L at 25 mM) and supplemented with 100 units/mL Penicillin, 50 mg/mL Streptomycin, and 10% FBS. The cells were maintained in a humid chamber at 37°C in an atmosphere of 5% CO2.
Cell Analysis
Variation of ALA solutions dose and time: Cells were cultured in 14 plates, and with 80% of confluence, the medium cells were discarded, the cells were washed with PBS and 1 mL of filtered ALA solution (in MilliQ water) of 200 μg/mL, pH7.4, was added to each well with 8 mL of culture medium DMEM serum-free (final ALA concentration of 0,13 mM/plate). For the control group (ALA0h) were added 8 mL of DMEM to the cells (without serum) with 1 mL of Milli Q water (without ALA). After 30 minutes, 1, 4, 8 and 12 hours the medium with ALA solution was removed, cells were washed, trypsinized and centrifuged. For dose ALA variation, to the cells were added from 0 to 0.5 mM of ALA to each well, and after 4h, the ALA solution was removed and the cells were trypsinized. To the cells pellets were added 1 mL of acetone, mixed into a vortex and centrifugedat4000 rpm for 15 min. The fluorescence analysis was carried out at the same day. All experiments were done in triplicate. Migration of PpIX from cells to blood: DU145 cells were cultured in 6-well cell culture plate. With 80% of confluence, the cells received the ALA solution (0.35 mM) and after four hours, the ALA solution was removed, the cells were washed with PBS solution several times to remove residual ALA, and then it was replaced by 300µL whole blood of healthy mice (collected from retro-orbital plexus 30 minutes before). All experiments were performed in accordance with the institutional guidelines animal care (approved by the UNIFESP Ethics Committee on animal Research -CEP 0667/08). The blood remained in contact with the cells for 2 hours.
ALA Solution
Solutions of ALA (Sigma) were freshly prepared by dissolving 0.045 g of ALA in 4.5 mL of phosphate buffered saline (PBS). The pH was adjusted to ~7 by addition of NaOH solution.
Porphyrin Extraction
The whole blood remained in contact with the cells for 2 hours, after this period it was removed and PpIX was extracted by the addition of 500µL grade acetone, centrifugation at 4000 rpm for 15 minutes and analyzed spectroscopically at the same day. The DU145 cells were washed with PBS solution to remove residual blood prior to tripsinization. Then, they were centrifuged and re- suspended in 500µL of analytical grade acetone. The mixture was centrifuged at 4000 rpm for 15 min. The clear supernatant of mixture was stored in a clean tube and spectro fluorometer analyses were carried out at the same day.
Fluorescent Spectral Analyses
The emission spectra were obtained by exciting the samples at 405 nm, inside of a 1 mm optical path cuvette. The fluorescence of the samples was analyzed with a Horiba Jobin Yvon Fluorolog 3 Fluorimeter in the range of 575 -725 nm.
Results
Initially, it has been studied ALA-induced PpIX fluorescence emission spectrum in DU145 cells, from 0 to 12 hours. Untreated DU145 cells present a very low PpIX signal intensity, (ALA 0h, Figure 1). However, the administration of ALA causes a time-dependent increasing in the concentration of PpIX into these cells. 12 hours after ALA administration the emission signal of PpIX was 200-fold higher than the samples without ALA (ALA 0h). PPIX emission spectrum comprises two major bands, one around 630 nm and another one around 700 nm.
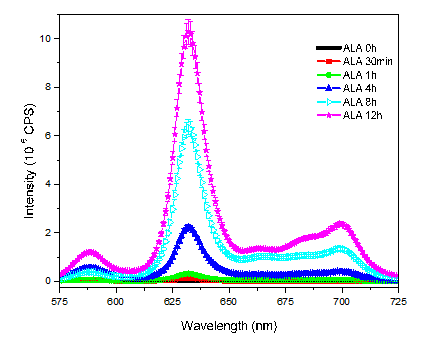
The production of PpIX by DU145 cells were analyzed at different doses from 0 to 0.5 mM/plate, pH 7.4, for a period of approximately 4 hours. Again, the 0 mM ALA group corresponds to the group that received just MilliQ water (without ALA).
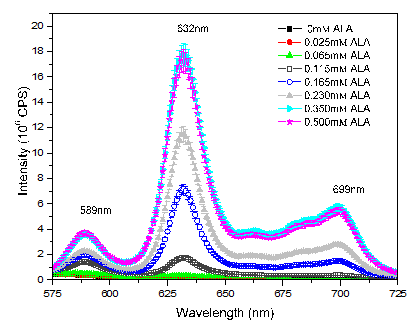
Figure 2: Mean fluorescence spectra of PpIX from DU145 cells exposed during 4h 30 to different concentration of ALA from 0 to 0.5mM/plate In this case, the cells showed an increasing of PpIX production until the concentration of 0.35 mM of ALA per plate. Thereafter, the production of PpIX remained unchanged, indicating that the cells have already exhausted their capacity to synthesize PpIX, probably due to the enzymes limitation.
Migration of Ppix from Tumor Cells to Blood
A series of experiments were carried out to study PpIX transference from the tumor cells to the blood cells. It is well know that when ALA is administrated to animals, significant levels of PPIX reaches a peak in few hours (approximately 3 hours), returning to background levels by 24 hours [13]. The extra PPIX synthesized reaches the blood [14] and is eliminated from the organism even by feces [15], since a higher concentration is toxic [11]. Although the excess of PPIX is prejudicial for the cells, in the in vitro experiments, the cells the cells retained the PPIX produced, at least for several hours see (Figure 1). It suggest that a mechanism to take it out from cells should exist. (Figure 3A) shows the mean of PPIX auto fluorescence emission spectrum of whole blood of healthy mice (N=5). Figure 3B represents the PPIX auto fluorescence emission extracted from DU145 cells cultured in DMEM medium. The weak fluorescence intensity signal of the band at 632 nm was 5.5 x104 CPS.As shown in Figure 3C, the contact of DU145 tumor cells with whole blood from healthy mice, for two hours, has not altered the intensity of porphyrin band of both samples. DU145 cells exhibit a small amount of endogenous PpIX to transfer it to the blood. In the following experiments, our aim was to verify if the DU145 tumor cells would transfer the extra PpIX synthesized, induced by ALA, from intracellular space to culture medium and/or to the blood. Herein, the cells received ALA solution and they were maintained in a humid chamber at 37°C in an atmosphere of 5% CO2 for 4 hours. In the first experiment, PpIX was extracted from the DU145 tumor cells as soon as the ALA solution was removed (Figure 3D). In the second one, ALA solution was replaced by DMEM medium and the cells were maintained in a humid chamber at 37°C in an atmosphere of 5% CO2 for 2 hours (Figure 3E). Figures 3D & 3E show the same emission intensity confirming that the excess of PpIX produced in the presence of ALA was not transferred to the DMEM medium, at least during the first two hours. Additionally, the cells ended the PpIX synthesis when ALA solution was removed. In this situation, the cells showed a mean emission intensity of 9.6 x 106 CPS, which represents an emission value 175- fold higher than the endogenous emission (Figure 3B). Finally, in the third case, after the incubation with ALA solution for 4h, the solution was removed, the cells were washed with PBS solution and 300μL of whole blood of healthy animals were added to the plate and they were maintained in a humid chamber at 37°C in an atmosphere of 5% CO2 for 2 hours. After this period, blood and cells had their PPIX extracted with acetone and the supernatant were analyzed, separately. The mean emission intensity of PpIX extracted from cells was 2.9 x 106 CPS, which represents only 30% of the PpIX synthesized in the presence of ALA, as seen in Figure 3.D and 3.E. While in the whole blood, just the opposite occurs, i.e., the emission signal was 4.9 x 106 CPS, an increased value of almost twice in comparison with Figure 3.A or 3.C (2.8 x 106 CPS). This data confirms that the PPIX concentration increasing in whole blood samples resulting from the transference of PpIX excess from intracellular space into the blood; and moreover it is not a simple diffusion process, but a complex efflux system that cells have developed to transport and regulate PpIX concentration (since an excess of PPIX could be highly toxic to the cell), what was confirmed by the transference of PPIX into blood, but not into DMEM medium. This result corroborates the literature reports [11, 12]. Another test performed was to add ALA into the blood of animals collected and incubated in a petri plate for two hours. There was no difference in the emission signal from the blood, since mature erythrocytes are unable to synthesize more PPIX (results not shown because it is the same spectrum of Figure 3A).
![Figure 3: D and 3.E. While in the whole blood, just the opposite occurs, i.e., the emission signal was 4.9 x 106 CPS, an increased value of almost twice in comparison with Figure 3.A or 3.C (2.8 x 106 CPS). This data confirms that the PPIX concentration increasing in whole blood samples resulting from the transference of PpIX excess from intracellular space into the blood; and moreover it is not a simple diffusion process, but a complex efflux system that cells have developed to transport and regulate PpIX concentration (since an excess of PPIX could be highly toxic to the cell), what was confirmed by the transference of PPIX into blood, but not into DMEM medium. This result corroborates the literature reports [11,12]. Another test performed was to add ALA into the blood of animals collected and incubated in a petri plate for two hours. There was no difference in the emission signal from the blood, since mature erythrocytes are unable to synthesize more PPIX (results not shown because it is the same spectrum of Figure 3A).](/fulltextimages/551/fig_3.png)
Conclusion
This study indicates that the excess of PpIX synthesized by tumor cells in the presence of ALA is transferred from them to the blood. Thus, it indicates that the higher PpIX concentration found in the blood of tumor animal models and cancer patients is probably produced by tumorigenic cells and then transferred to blood flow through an efflux system, not just by a simple diffusion process.
Acknowledgment
The authors acknowledge the Brazilian funding agency CAPES for the financial support. The authors declare that there is no conflict of interest.
References
-
Fotinos N, Campo MA, Popowycz F, Gurny R, Lange N (2005) 5-AminolevuIinic Acid Derivatives in Photomedicine: Characteristics, Application and Perspectives. Photochem Photobiol 82(4): 994-1015.
-
Abels C, Heil P, Dellian M, Kuhnle GE, Baumgartner R, et al. (1994) In vivo kinetics and spectra of 5- aminolaevulinic acid induced fluorescence in an amelanotic melanoma of the hamster. Br J Cancer 70(5): 826-833.
-
Kriegmeir M, Baumgartner R, Kneuchel R, Stepp H, Hofsteder F, et al. (1996) Detection of early bladder cancer by 5-aminolevulinic acid induced porphyrin fluorescence. J Urol 155(1): 105-110.
-
Otake M, Nishiwaki M, Kobayashi Y, Baba S, Kohno E, et al. (2003) Selective accumulation of ALA-induced PpIX and photodynamic effect in chemically induced hepatocellular carcinoma. Br J Cancer 89(4): 730-736.
-
Gibbs-Strauss SL, O'Hara JA, Hoopes PJ, Hasan T, Pogue BW (2009) Noninvasive measurement of aminolevulinic acid-induced protoporphyrin IX fluorescence allowing detection of murine glioma in vivo. J Biomed Opt 14(1): 014007.
-
Larsen ELP, Randeberg LL, Gederaas OA, Arum CJ, Hjelde A (2008) Journal of Biomedical Optics 13.
-
Utsuki S, Miyoshi N, Oka H, Miyajima Y, Shimizu S, et al. (2007) Fluorescence-guided resection of metastatic brain tumors using a 5-aminolevulinic acid-induced protoporphyrin IX: pathological study. Brain Tumor Pathology 24(2): 53-55.
-
Silva FRO, Bellini MH, Tristão VR, Schor N, Vieira ND (2010) Intrinsic Fluorescence of Protoporphyrin IX from Blood Samples Can Yield information on the growth of prostate tumours. Journal of Fluorescence 20(6): 1159-1165.
-
Kalaivani R, Masilamani V, Sivaji K, Elangovan M, Selvaraj V, et al. (2008) Photomedicine and Laser Surgery 26(3): 251-256.
-
Masilamani V, Al-Zhrani K, Al-Salhi M, Al-Diab A, Al- Ageily M (2004) Cancer diagnosis by autofluorescence of blood components. J Luminescence 109(3-4): 143-154.
-
Krishnamurthy P, Xie T, Schuetz JD (2007) The role of transporters in cellular heme and porphyrin homeostasis. Pharmacology & Therapeutics 114(3): 345-358.
-
Quigley JG, Yang Z, Worthington MT, Phillips JD, Sabo KM, et al. (2004) Identification of a Human Heme Exporter that Is Essential for Erythropoiesis. Cell 118(6): 757-766.
-
Chang SC, Buonaccorsi GA, Mac Robert AJ, Bown SG (1997) Interstitial photodynamic therapy in the canine prostate with disulfonated aluminum phthalocyanine and 5-aminolevulic acid induced protoporphyrin IX. Prostate 32 (2): 89-98.
-
Silva FRO, Nabeshima CT, Bellini MH, Nabeshima, Schor N, et al. (2011) Enhancement of blood porphyrin emission intensity with aminolevulic acid administration: A new concept for photodynamic diagnosis of early prostate cancer. Photodiganosis and Photodynamic Therapy 8(1): 7-13.
-
Silva FRO, Nabeshima CT, Bellini MH, Schor N, Vieira ND Jr, et al. (2013) Study of Protoporphyrin IX Elimination by Body Excreta: A New Noninvasive Cancer Diagnosis Method. J Fluoresc 23(1): 131-135.
- Cancer Diagnosis from RNA Sequence of Blood Cells by Using AI
- Field Cancerization in Oral Cavity, Case Report and Review of Literature. Oncologic Program Salud Integral Hospital, Managua, Nicaragua
- Identification of B Lymphocytes in Cancer Patient’s Blood
- A Case Report of a Breast Cancer Patient Developing Pneumonitis as a Result of Abemaciclib Therapy
- Immune Checkpoint Therapeutics for Today’s Fight and Beyond
- The Amalgamated Sophomore-Gonadoblastoma