A Review of Endophytic Fungi and their Applications in Different Fields of Biotechnology
Endophytic fungus is an intriguing group of organisms that live inside their host's living tissues, which are primarily higher plants. Endophytes generate naturally occurring bioactive chemicals that are thought to operate as an elicitor for the synthesis of secondary metabolites in plants, without causing any disease symptoms in the host cells. By imparting stress tolerance and enhancing host resistance to a range of diseases, this incredibly varied collection of fungus can have a significant influence on plant ecosystems. They are known to improve nutrition through the reversible exchange of nutrients and protect plants from infections, which in turn affects development by releasing plant growth hormones. Endophytic fungus-infected plants exhibit notable increases in biomass, enhance commercial plant output, and are therefore beneficial to the agricultural industry. The potential uses of endophytic fungi as biological vectors, biological control agents, and insecticidal products, sources of secondary metabolites, antimicrobial agents, antitumor compounds, antibiotics, immunosuppressants, and antidiabetic agents make them of biotechnological interest. The current study concentrated on the numerous uses of fungal endophytes in several biotechnology fields.
Introduction
Endophytic microorganisms, such as bacteria and fungi, are ubiquitous in all plant species and inhabit the internal tissues of the plant without causing any apparent detrimental effects [1]. Endophytic microorganisms that promote plant growth (PGPE) reside within the tissues of plants, and their robust symbiotic relationship enhances the exchange of nutrients and enzymatic activity. Microbial endophytes can infiltrate plant tissues in a latent manner, thereby exhibiting no discernible symptoms [2]. Endophytic
fungi are a potential source of biocontrol agents by research that has been conducted on them [3]. These organisms spend part or all their life cycles living inside the otherwise healthy internal plant tissues of their hosts, yet they do not seem to do any noticeable damage to those tissues. They have a significant impact on the physiological processes that occur inside their host plants. By creating a diverse array of fungal metabolites, fungal endophytes improve their host’s tolerance to abiotic stress, disease, insects, and mammalian herbivores [4].
Endophytes may either directly or indirectly activate these plant growth-promoting features in plants [5]. Endophytes are also known to have antimicrobial properties. These fungi frequently play a role in nutrient intake, boosting plant growth, improving plants’ ability to withstand stress, limiting the growth of rivals and soil-borne pathogenic organisms, and improving disease resistance. Several techniques must be used in sustainable agriculture to improve food production while minimizing damage to the crop, the environment, and public health. An effective substitute for conventional chemical farming is the use of fungal endophytes to enhance plant development. When crops are exposed to harmful biotic and abiotic stress factors including drought, cold, salinity, and disease, poisonous and lethal chemical pesticides and fertilizers, endophytic fungi help the plant to cope up with the stress [6]. An alternative to conventional chemical plant growth boosters is the use of endophytic fungi. Using mechanisms including triggered systemic resistance, bioremediation, and biological control, endophytic fungi help plants more effectively endure the impacts of abiotic stress [3].
Applications of Fungal Endophytes
Beneficial secondary metabolites have been isolated from endophytic fungi, which have been used in biotechnological breakthroughs in the medicine, agricultural, nutrition, and environmental processing industries. Microorganisms called endophytes reside inside plant and are essential to their growth and well-being (Figure 1).
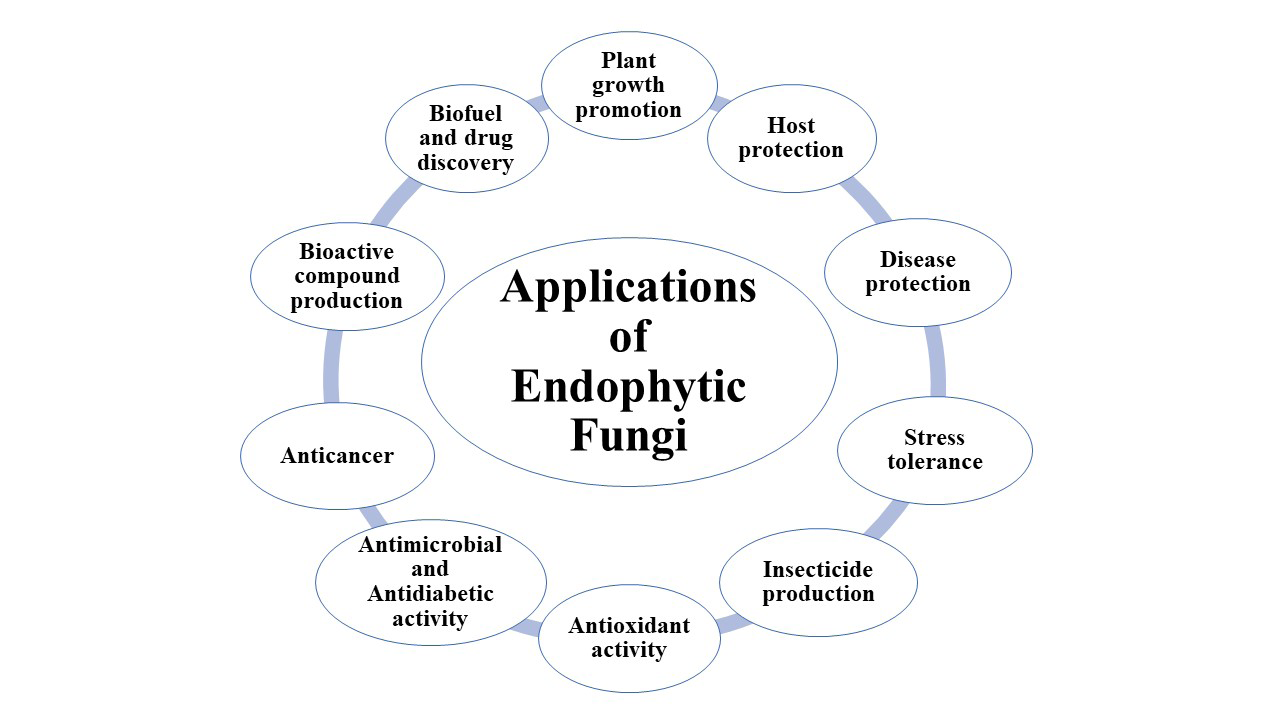
Protection of the Host Plant Against Herbivorous Animals
There has been a significant amount of research and literature on the extremely specialised nature of the symbiotic relationship between plants and endophytes, as well as the impact that fungal alkaloids have on both vertebrate and invertebrate herbivores. When herbivores consume plant tissue, they are certain to come into contact with fungal metabolites if the host tissue has been pervasively and systemically colonised with endophyte hyphae. Herbivores may range from huge animals to tiny arthropods. Insect resistance is increased by the majority of type C endophytes. Plants’ resistance to insect herbivory is shown to depend on both the kind of fungal strain and the stage of growth, as determined by Rashid and Chung [7]. Evidence of the antinematode activity of class 1 endophytes has also been given by other investigations [8]. Although many endophytes are thought to confer resistance to insects and nematodes on their host plants, studies have shown that this is not always the case. These findings emphasise the need to investigate endophytes in their natural habitats among native plant species [9]. While C-endophytes have traditionally been regarded as beneficial mutualistic organisms, recent research has revealed that a significant number of endophytes do not confer any protective advantages to their host plants [10]. Class IV endophytes, according to some studies, help the host plant defend itself from herbivores by secreting secondary compounds.
Plants with endophytes have been shown to be more resistant to fungal infection, and herbivores that nourish on infected plants have lower productivity. Endophytic fungi mitigate the negative effects of insect herbivores through a variety of mechanisms, such as diminished feeding, slowed development, stunted population growth and mortality [11]. Vertebrate herbivores exhibit similar avoidance and diminished performance patterns, including birds, rabbits, and deer. Endophyte infection can even lessen the impact of herbivores that live below ground, like nematodes and root-feeding insects. The strongest example for the anti- herbivore advantages of endophytic fungi comes from studies that indicate herbivore populations are destroyed when permitted to graze solely on plants infected with fungal endophytes [12].
Endophytic fungi create a wide variety of chemical defences, but not all of them are equally efficient, and many insect herbivores develop immunity to a specific substance at one point or another in their life cycles. As a general rule, larvae are more susceptible to poisons than adults. Geographic variation in consumption patterns means that even endophytic fungi that are supposed to be protective for their hosts, like Neotyphidium in plants, may not always cause avoidance or negative effects on herbivores [13].
Influence of Plant’s Ability to Withstand Stress
Plants are encountered with increasingly hostile environments for growth and maintenance as a result of environmental deterioration caused by agricultural operations and the changing climate. Equally difficult for a crop is that ever-increasing demands for agricultural output just further exacerbate the issue. Endophytic fungi are a potential solution for protecting plants from different stresses, and in this case, assistance is clearly required for healthy plant development. Endophytic fungi can protect host plants from water scarcity, extreme heat or cold, high or low salinity, and even poisonous metals [14]. Several dysfunctions in plant structure and function may be traced back to the altered genetic regulation of cellular pathways that results from exposure to abiotic stresses [15]. Endophytic fungus provides several benefits to host plants in a number of ways that increase their chances of success in the face of adversity. In response to oxidative stress, plants raise their catalase and peroxidase activity, which generates reactive oxygen species and initiates membrane damage and lipid peroxidation.
Endophytic fungi increase resistance to ROS and decrease lipid peroxidation [16]. Growth of the plants can be initiated directly by phytohormones, and indirect plant benefits can be achieved through the phytohormones’ role in modulating the adaptation process to abiotic stresses. The hormone abscisic acid regulates stress-response gene expression and stomatal closure, both of which serve to decrease transpiration and so fluid loss. ABA is suppressed by endophytic fungi’s presence [17]. Khan, et al. [18] investigated the effects of inoculating saline-stressed cucumber plants with the endophytic fungus Paecilomyces spp. Vegetative growth was higher in inoculation salinity-adapted plants. The favourable endophytic interaction of Yarrowia lipolytica was also reported by Jan, et al. [19], which reduced the negative effects of salt on maize plants. The effectiveness of the endophytic fungus Porostereum spadiceum in alleviating salt stress and enhancing the growth of soybean plants was observed. To achieve this, the phytohormone levels of both inoculated and control seedlings were compared [20]. Through regulating phytohormones, endophytic colonisation mitigated the negative effects of salt. The endophytic fungus Aspergillus niger was shown to give sunflower and soybean with resistance to high temperatures in another investigation. Plant length and biomass were all significantly raised by fungal inoculation [21]. Fungal endophytes produce bioactive metabolites and activate systemic resistance as the primary defence against pathogens, herbivores, and nematodes [22].
Because of its protective benefits on plants against biotic stresses, endophytic colonisation by a variety of fungi has been the centre of a significant amount of study. Several of these studies focused on Serependita indica, an endophyte that can protect many different types of crops from many different diseases [23]. Endophytic colonisation of cotton plants by Phialemonium sp., as described in a 2018 study by Zhou, et al. [24] inhibited root penetration by Meloidogyne incognita worms, gall development, and nematode reproduction. Endophytic colonization’s impact on plant development and resistance to pests has also been investigated. One study by Dash, et al. [25] used an inoculation of endophytic fungi. The objective of the study was to investigate the impact of endophytic invasion on the fitness of the host plant and its resistance against Tetranychus urticae, commonly known as the two-spotted spider mite. To achieve this, seeds of Phaseolus vulgaris were subjected to inoculation with Bacillus thuringiensis, Isaria fumosorosea, and Lecanicillium sp. All strains examined by the scientists were able to successfully colonise bean plants as endophytes, and they were retrieved from the different parts of plants. The length of the plants and the fresh weight of the shoots and roots were both improved by the treatment of the seeds. Endophytic associations between Penicillium sp.
and Phoma glomerata, found in cucumber plants, resulted in considerably greater biomass and enhanced growth despite salt and water stress. The friendly relationship enhanced potassium, calcium, and magnesium intake while reducing sodium toxicity during saline stress. Reprogramming of plant growth in response to abiotic challenges by fungi via regulation of abscisic, jasmonic, and salicylic acid production has also been identified [26]. Studies by Jaber and Enkerli [27] demonstrated the ability of various strains of B. bassiana and M. brunneum to establish fungal colonies in broad bean plants, resulting in increased plant growth in seed-treated individuals. The same species were later reported by Jaber [28] as having successfully colonized the wheat (T. aestivum) plant’s aerial parts and roots to stimulate plant development. Due of the impressive ability of the fungus to regenerate from both leaves and stems, suggested the capacity of systemic colonization by M. endophytica, which was identified in 91% of the plants grown from fungal-inoculated seeds. These researches emphasized the possibility of endophytic interactions between several fungal species and their hosts. Research into the relationships between helpful microbes and their plant hosts has led to the discovery of plant- microbe interactions, opening up a new avenue in the quest for environmentally friendly farming practices [29].
Production of Insecticidal Agents by Endophytes
Many endophytic fungi have anti-insect properties and are proved active against insects by producing some toxic insect repellent substances, some of which are given below in Table 1.
| Sr. No. | Producing Fungus | Compound | References |
|---|---|---|---|
| 1 | Geotrichiumcandidium | Neofrapeptin A | Fredenhagen, et al. [30] |
| 2 | Cordyceps heteropodac | Cicadapeptin I | Krasnoff, et al. [31] |
| 3 | Omphalotusolearius | Omphalontins E-I | Liermann, et al. [32] |
| 4 | Fusarium sp. | Apicidin | Singh, et al. [33] |
| 5 | Penicillium citrinum | Quinolactide | Abe, et al. [34] |
| 6 | Aspergillus niger | Nafuredin- γ | Ōmura, et al. [35] |
| 7 | Beauveria bassiana | Beauverolide N | Kuzma, et al. [36] |
Table 1: Insecticidal compounds produced by endophytic fungi.
Plant Growth Promotion
Endophytes, in addition to coping with host defence, are required to release compounds that are antibacterial and antifungal to maintain a healthy balance of antagonism with other competing organisms. The synthesis of phytohormones, which encourage the development of plants, is an additional advantage of this process. Endophytes can indirectly or directly increase plant growth and yield (through biological nitrogen fixation, phytohormone production, siderophore synthesis, and phosphate solubilization), or indirectly through a variety of mechanisms (Table 2). Endophytes also have the potential to remove soil contaminants through the enhancement of phytoremediation (Figure 2).
| Sr. no. | Activity Exhibited by Endophytic Fungi | References |
|---|---|---|
| 1 | IAA production | Bric, et al. [37] |
| 2 | Phosphate solubilization | Talukdar, et al. [38] |
| 3 | Cellulolytic activity | Dar, et al. [39] |
| 4 | Ammonia detection | Szilagyi-Zecchin, et al. [40] |
| 5 | HCN production | Donate-Correa, et al. [41] |
| 6 | Amylase production | Hankin, et al. [42] |
| 7 | Siderophore production | Schwyn, et al. [43] |
Table 2: Different types of plant growth promoting activities exhibited by endophytic fungi.
![Figure 2: Plant growth promotions by endophytic fungi in different ways [44].](/fulltextimages/12812/fig_2.png)
Several fungal endophytes can stimulate growth in the host plant either directly or indirectly. This is achieved by processes such as habitat restriction and competition, direct antagonism of pathogens, and changes in the levels of plant hormones such as gibberellins and auxin in the host plant. The term “major nutrient” usually refers to this substance. It keeps plants in good overall health and vigour by doing things like encouraging the growth of roots, boosting the strength of stalks and stems, encouraging the production of flowers, improving crop quality, and boosting the resistance of plants to illnesses. Phosphorus is an important component of ATP. It is an essential component in passing on information about a plant’s genetic makeup from one generation to the next, acting as blueprint that details all aspects of a plant’s development as well as any changes to its DNA or RNA [45].
Because phosphorus is essential to the growth of seeds, it is abundant in seeds and fruits. Phosphorus is also present in high concentrations in fruits. Phytin is the primary type of phosphorus storage that is found in seeds. Phosphorus deficiency causes plants to mature later, produce lesser quality fodder, fruits, vegetables, and grains, and be more susceptible to disease; it slows the growth of the stem more than that of the roots and causes a purple hue to emerge in the leaves of some plants [46].
The organic form and the inorganic form of phosphorus are the two categories that may be found in the ground. It may be found in its organic form in decomposing plant matter, compost, and the tissues of microbes [47]. Plants are unable to make use of phosphorus in an insoluble state because it cannot be dissolved in water [48]. Fungi are the organisms most capable of releasing vital nutrients from soil, according to a great number of studies. It was found that Penicillium and Aspergillus were two key species that were discovered with strong mineral-solubilizing activity, and this activity was associated to a lowering in the pH of culture medium.
Inorganic phosphate salts break down because of microorganisms, which leads to the release of organic acids [49, 50]. Sustainable agriculture benefits from the wide variety of fungi that are known to reside in endophytic interactions with plants and are generally considered helpful to plant growth and development. Penicillium sp. was isolated from tea leaves, and their phosphate-solubilizing activity was investigated by Nath, et al. Both isolates that have demonstrated exceptionally high phosphate solubilization activity have contributed to a gradual elevation in the acidity of the culture medium up to day 8. These isolates may find widespread use as biofertilizers. Isolation of Trichoderma gamsii from Lens esculenta lateral root endophytes was confirmed by microscopic analysis, morphological analysis, and 18S rDNA sequencing. With a drop in pH, this fungus in the growth medium was able to solubilize up to 17% of the phosphate. Trichoderma gamsii demonstrated its capacity to promote plant development in a greenhouse bioassay utilizing four different test crops [51]. Research that was carried out by Nath, et al. [52] focuses on isolating fungal endophytes from tea (Camellia sinensis) roots, stems, and leaves that were collected from tea gardens in Assam, India. These endophytes were then subjected to in vitro testing to determine their PGP activity levels. The fungus Aspergillus niger was shown to have the highest level of IAA activity, followed by the fungus Penicillium sclerotiorum. The highest activity was demonstrated by Fusarium oxysporum followed by Penicillium chrysogenum F1. Penicillium sclerotiorum is the most efficient phosphate solubilizer. Phytohormones are natural compounds that are released in certain organs of plants. These substances can be transported to other locations, where they stimulate specific reactions in terms of the plant’s biology, physiology, and morphology. Aside from that, plant microbiomes synthesize growth regulators including gibberellic acid, auxins, and cytokinin [50]. The microbiomes of plants have been found to play a significant role in the development and proliferation of various plant organs, including but not limited to flowers, stems, leaves, and fruits.
Plant growth hormones can enhance and actually impact cell growth, development, and differentiation even at low concentrations. The following are the five major classes of phytohormones: abscisic acid (a) auxins, (b) cytokinins, (c) ethylene, (d) Gibberellins, (e) abscisic all are important plant growth regulators. According to the research conducted by Hamayun, et al. [53], certain fungal endophytes facilitate the growth of their host by stimulating the synthesis of gibberellins (GAs), indole-3-acetic acid (IAA), and cytokinins. The endophytic fungus, Cladosporium sphaerospermum, was identified in the roots of Glycine max (L.) and has been demonstrated to produce gibberellins. Rice was able to flourish when it was exposed to the culture filtrate, which had higher concentrations of the growth factors GA3, GA4, and GA7 respectively. The roots of Calystegia soldanella were examined, and a fungal endophyte called Cadophora malorum that produces GAs was discovered [54]. RSF-4L and RSF-6L are fungal endophyte species that have been identified as belonging to the genus Fusarium and a species of Alternaria were isolated from the leaves of the Solanum nigrum plant. According to the results of the Salkowski experiments, both Fusarium sp. &Alternaria sp. produced IAA, respectively. After being treated with fungal CFs, plant growth characteristics were enhanced.
The two categories of fungal endophytes play a crucial role in facilitating the growth and development of plants, as noted by Khan, et al. [55]. The presence of fungal endophytes significantly enhances a plant’s capacity to endure biotic and abiotic stresses. To aid in relationships with plant hosts and to provide resistance to pests and diseases, endophytic fungi produce bioactive compounds. These bonds are mediated and protected by bioactive chemicals. There is enough proof to demonstrate that endophytic fungi interact with their hosts for mutual advantages, including the improvement of host health, defence against pests, grazers, and rhizosphere nematodes, and improvement in drought tolerance and root development. In exchange, endophytic fungi acquire habitation and diet.
Endophytes are microorganisms that synthesise bioactive compounds, enabling plants to defend themselves and providing them with the requisite energy to do so. Endophytic microorganisms create mycotoxin, which defends the host plant against attack by pathogenic microbes caused by the plant’s natural predators [56]. The 18S rRNA gene sequencing shows that the strain belonged to the genus Penicillium and produced gibberellins with the GA5 designation [53]. Fungal extracellular enzymes are finding more and more applications in fields as diverse as biotechnology, medicines, the food and beverage industries, leather production, agriculture, bioremediation, and chemical compound biotransformation [57, 58].
In addition to their role as phytoremediators, endophytes perform the functions of main saprobic decomposers, mutualists, and latent pathogens. There is a wide variety of endophytes that can perform the function of phytoremediation. The majority of these endophytes are heavy metal-resistant endophytes, both hyper-accumulators and non-hyperaccumulators and organic contaminant- degrading endophytes [59, 60]. Bioremediation using these endophytes increases plant growth (via regulation of phytohormones [61], enzyme production, and nitrogen fixation), decreases phytotoxicity (via production of iron chelators, siderophores, and enzymes that degrade iron), and ultimately increases plant metal tolerance [62].
Antimicrobial and Antidiabetic Activity
Endophytes are capable of producing several hydrolytic enzymes in addition to peptide and polyene macrolide antibiotics like amphotericin B and nystatin [63]. Penicillin and cephalosporins are, without a doubt, the most significant anti-infective medications now available on the market. Endophytic fungi produce a number of antibiotics, including Daptomycin [64] and Valinomycin, a strong antibiotic that is effective against acute respiratory syndrome Coronavirus [65, 66]. An endophytic fungus Cryptosporiopsis quercina is renowned for its active antifungal action against C. albicans and other species of Trichophyton. These fungi are considered to be human fungal infections. A molecule known as cryptocandin [66], which was discovered from C. quercina, is renowned for its efficacy as an antimycotic drug. A compound called cryptocin was isolated from the same fungus and has been demonstrated to effectively combat Pyricularia oryzae [67].
Pseudomycins, also known as lipopeptides, have been shown to be active action against human pathogenic fungi such as Candida albicans and Cryptococcus neoformans [68, 69]. In order to develop NMR approaches, ambuic acid, which is generated by Pestalotiopsis microspore, in conjunction with another endophyte called terrein, is employed [70]. A molecule called pestaloside [71], which was isolated from P. microspore, has been shown to have antimicrobial characteristics, but the two sesquiterpenes known as pestalotiopsins A and B are known to have phytotoxic effects. Muscodor albus, in addition to endophytic fungus, is responsible for the production of volatile chemicals that have antimicrobial and antifungal properties [72]. The isoamyl acetate that is generated by this fungus is the most useful substance that it produces. There is evidence that some species of Gliocladium generate the volatile antibiotic chemical annulene [73]. Some bioactive agents produced by the genus Xylaria, such as sordaricin and multiplolides A and B, have been shown to have biological activity against C. albicans [74]. On the other hand, mellisol and 1,8-dihydroxynapthol 1-O-a-glucopyonaside have been shown to have activity against HSV type 1 [75]. An endophytic Streptomyces species that was isolated from Grevillea pteridifolia generates a new antibiotic known as ‘Kakadumycin A,’ which is both an effective antibacterial agent and an antimalarial agent [76].
The antimicrobial spectrum of Phomopsis sp., which was isolated from the host plant Rhizophora apiculata, was more effective against gram-positive bacteria [77]. D. zingiberensis culture filtrates inhibited Bacillus, Staphylococcus and E. Coli [78]. It was discovered that the fungal endophytes found in Eucalyptus exserta have strong antibacterial effects when tested as raw extracts [79].
Anticancer Compounds Produced by Endophytes
It is possible for endophytes to produce secondary metabolic products that are identical to or equivalent to those generated by their hosts (Table 3). Anticancer agents are examples of the bioactive substances that are created simultaneously by plants and the endophytes that are linked with them. Endophytic fungi have been shown to be a rich and consistent source of naturally occurring chemicals that have anticancer properties. These natural products have the potential to have a significant effect on the development of anticancer medications in contemporary medicine [80]. Endophyte-derived natural compounds have the potential to be employed as an alternative source in the research that ultimately leads to the creation of innovative treatment options for cancer [81, 82, 83]. Many endophytes have been identified as the sources of new chemicals that have shown promising results in anticancer testing.
| Sr. No. | Host plant | Fungal Endophyte | Anticancer Agent | References |
|---|---|---|---|---|
| 1 | Taxus brevifolia | Taxomyces andreanae | Taxol (Palcitaxel) | Stierle, et al. [84] |
| 2 | Camptotheca acuminate | Fusarium solani | Camphothecin | Ran, et al. [85] |
| 3 | Podophyllum. Hexandrum | Tramete shirsuta | Podophyllotoxin | Giri, et al. [86] |
| 4 | Adenophoreaxiliflora | Chaetomium sp. | Chaetominine | Jiao, et al. [87] |
| 5 | Torreya. Taxifolia | Pestalotiopsis microspore | Cytochalasins Torreyanic acid | Sappapan, et al. [88] |
Table 3: _Anticancer agents produced by fungal endophytes._
Bioactive Substances Produced by Endophytic Fungi
Interaction of plant with the endophytic fungus naturally results in the biosynthesis of secondary metabolites (Table 4). Genes from both the plant and the endophyte species regulate and modulate the nature of the connection between them [89]. Endophytic fungi in medicinal plants produce an abundance of secondary metabolism products [88]. Secondary metabolite production in plants may also be influenced by the symbiotic interaction between endophytes and their host plants. Plants developing in adverse environments must be tested for endophyte isolation and its metabolites [88]. Alkaloids, benzopyranones, chinones, flavonoids, phenolic acids, and many others are only some of the beneficial secondary metabolites that endophytes produce [90]. Such bioactive metabolites are commonly used as agrochemicals, antibiotics, antiparasitics and antioxidants [88]. Biotechnological methods that employ certain organisms show promise as viable substitutes for developing an infinite, low-cost, and sustainable source of high-quality bioactive products as well as aromatic compounds.
| Bioactive Compound | Plant Species | Endophyte | References |
|---|---|---|---|
| Podophyllotoxin and its analogues | Sinopodophyllum hexandrum, | Alternaria sp. | Giri, et al. [86] |
| Diphylleia sinensis | Penicillium sp. | ||
| Dysosmaveitchii | Monilia sp. | ||
| Camptothecin and its analogues | Camptotheca acuminata, | Fusarium solani | Kusari, et al. [91] |
| Nothapodytes foetida | Botryosphaeria parva | ||
| Huperzine A | Huperzia serrata | Acremonium | Kusari, et al. [91] |
| Lycopodium serratum | Penicillium chrysogenum | ||
| Vinblastine and its analogues | Catharanthus roseus | Alternaria sp. | Huang, et al. [92] |
| Cytoskyrin a | Conocarpus erecta | Cytospora sp. | Brady, et al. [73] |
| Phomoxanthone a | Lucila spinosa | Xylariasp. | Isaka, et al. [93] |
| Phomoxanthone b | Tectona grandis | Phomosissp. | |
| Rubrofusarin b | Cynodondactylon | Aspergillus niger | Guo, et al. [94] |
| Emindole DA | Mediterranean green alga | Emericellanidulans | Kralj, et al. [95] |
| Paclitaxel and its analogues | Taxus cuspidata | Alternaria sp. | Wani, et al. [95] |
Table 4: Bioactive compounds produced by Endophytic Fungi.
Production of Antioxidants
ROS, or reactive oxygen species, are by-products of the aerobic process found in all living organisms that serve a number of functions. The production of free radicals in the form of various ROS is triggered when the light absorption capacity of photosynthetic cells exceeds their usage [97, 98]. This happens because oxygen molecules can quickly accept electrons from the body’s electron transport system [99]. Herbivorous and pathogenic plant stress causes the production of ROS via oxidative bursts in the host plant [100, 101]. During these bursts, chemicals are synthesised that may be employed to either block the spread of invading hyphae physically or to have direct, damaging effects on the cells of these organisms. Many studies [102, 103] corroborate this notion. This study’s results provide credence to the hypothesis that reactive oxygen species serve a useful role in preparing the plant’s response to abiotic stresses, which has been proposed after a thorough revaluation of the role of ROS in the abiotic stress response of plants [99, 104]. The plant produces many singlet oxygen species as part of this function, and these species travel extensive distances inside the tissue of the plant to provide systemic communication throughout the plant.
An arsenal of antioxidants is produced during the activation of the plant’s stress response. These antioxidants then regulate the degree of ROS build-up in plant cells, hence lowering the risk of cell damage and apoptosis [101]. ROS have a mechanistic role in apoptosis, nonspecific physiological stress, and systemic signalling, all of which influence the effectiveness of fungal infection or endophytic fungi colonisation and the plant’s reactions, which may be resistance, acceptance, or sanctions. In addition, via the interaction between oxidants and antioxidants, antioxidants may play a role in the transmission of stress signals. As a result, the host may be better able to respond rapidly to pathogenesis and tell the difference between a mutualist and a pathogen through chemical communication with an avirulent pathogen or an asymptomatic endophyte.
When cultures of Phyllosticta sp. were subjected to ROS, Srinivasan_,_ et al. found that the organisms exhibited a high level of antioxidant activity. The interaction between fungal endophytes and host plants is a dynamic process that involves the potential involvement of reactive oxygen species (ROS) and antioxidants in the modulation of the host’s hypersensitivity and systemic developed resistance mechanisms.
Biofuel Production
Researchers looked at the possibility of producing lipid biodiesel precursors using endophytic fungus that had been identified from a variety of different tropical plant species. Widespread interest is already being shown in the production of next-generation fuels using endophytic fungus as a hitherto identified reservoir of low - molecular weight hydrocarbons and lipids. These microbes will be directly responsible for the production of these forthcoming biofuels, which will result in a simpler and shorter biosynthetic process. In addition, they will be well suited for instant use in current combustion engines without requiring fundamental modifications to vehicle structure. Endophytes that have optimal lipid profiles have been reported to have been isolated by research organisations operating in different parts of the globe. These endophytes may be used as precursors in the production of biofuels. Researchers have discovered that some types of fungal endophytes may create volatile organic molecules, which can then be used to make traditional diesel fuel. The genomic sequence of one of these endophytic fungi that produces biofuel was only published not too long ago. Understanding of the biosynthetic mechanism by which endophytes are able to accomplish the difficult task of producing volatile hydrocarbons will be improved thanks to the discovery of Ascocorynesarcoides. Research in this vitally essential subject of alternative bio-energy for the purpose of ensuring India’s energy supply has gradually begun to acquire pace. Fungal endophytes live inside plants and are used in the production of biodiesel. The plants J_atropha_ curcas, Pongamia pinnata, Sapindusmukorossi, Mesuaferrea, Terminalia bellerica, Casabelathevetia, and Ricinus communis are some of the plants that contain these fungi [105].
Discovery of New Drugs
Natural substances have been shown in recent research to have potential in the pharmaceutical industry as a source of innovative and diversified pharmacological templates for future drug development and discovery [106, 107, 108, 109]. Research into endophytes for natural products is expected to play a significant role in the future of drug discovery and development since it is becoming increasingly clear that many natural compounds result from microbial associations with their hosts [110, 111].
Natural Products Derived from Fungi Endophytes
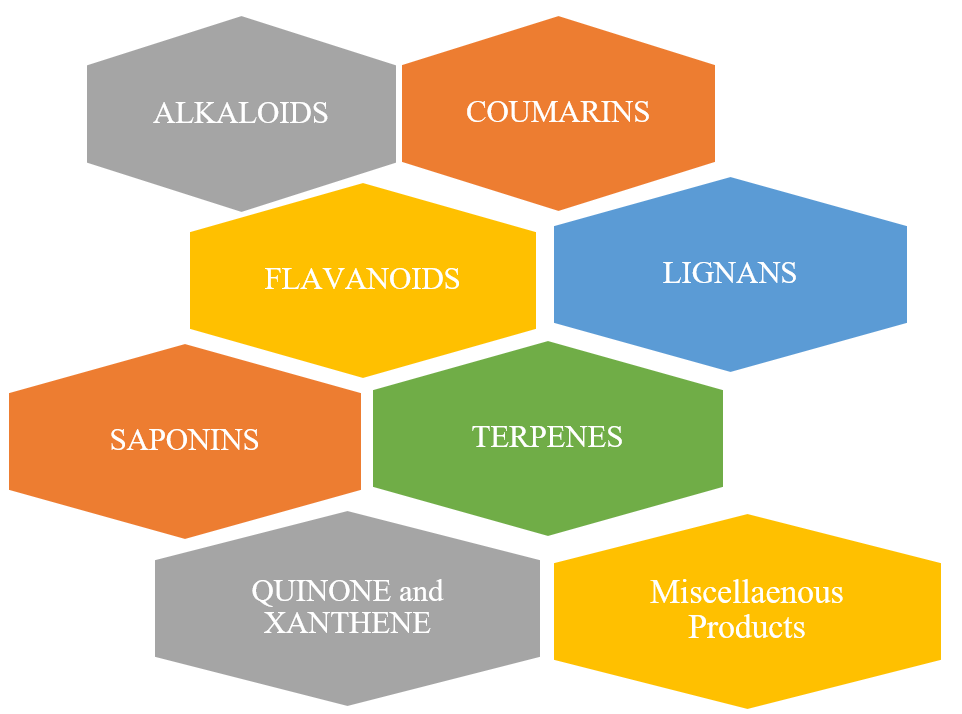
Endophytes of fungi are well-established sources of natural compounds and produce particular plant-derived metabolites that are beneficial from a pharmaceutical standpoint. Many kinds of plant-derived medicinal chemicals (Figure 3), which are generated by fungal endophytes, are discussed in Table 5.
| Endophytic Fungus | Plant Source | Plant Derived Compound | Activity | References |
|---|---|---|---|---|
| Alkaloids | ||||
| Cladosporium cladosporioides | Aconitum spp. | Aconitine | Anticancer | Yang, et al. [112] |
| Alternaria alternata | Capsicum annuum | Capsaicin | Anti-inflammatory, gastro-stimulatory | Devari, et al. [113] |
| Fusarium oxysporum, | Amoora rohituka Dysoxylum binectariferum | Rohitukine | Anticancer, CDK inhibitor | Kumara, et al. [114] |
| Coumarins | ||||
| Botryodia plodiatheobromae | Citrus bergamia, Grapefruit peel | Bergapten, Meranzin | Antioxidant | Zaher, et al. [115] |
| Annulohypoxylona bovei var. microspora | Acanthopanax senticosus, Sarcandra glabra | Isofraxidin | Anticancer | Yamazaki, et al. [116]; Cheng, et al. [117] |
| Penicillium sp., Xylaria sp. | Alibertia macrophylla | Mellein | Antibacterial, antifungal | Oliveira, et al. [118] |
| Penicillium sp. | Artemisia scoparia, Scopolia carniolica | Scopoletin, Umbelliferone | Antifungal, antioxidant, anti-inflammatory | Huang, et al. [92] |
| Flavanoids | ||||
| Colletotrichum sp | Cajanus cajan | Apigenin | Antibacterial, anticancer, antioxidant, | Shukla, et al. [119] |
| Hypocrealixii | Cajanus cajan | Cajanol | Anticancer, antimicrobial | Zhao, et al. [120] |
| Curcuma wenyujin | Curcuma spp | Curcumin | Anti-inflammatory, antioxidant, antitumor | Wang, et al. [121] |
| Lignans | ||||
| Members of xylariaceae | Angelica archangelica | Coniferin | Antidiabetic | Chapela, et al. [122] |
| Alternaria neesex | Forsythia suspensa | Podo-phyllotoxin | Antitumor, antivirus | Kong, et al. [123] |
| Saponins | ||||
| Penicillium oxalicum | Gymnema sylvestre | Gymnemagenin | Anti-diabetic | Parthasarathy, et al. [124] |
| Aspergillus sp. | Panax | Ginsenoside | antioxidation, antitumor | Wu, et al. [125] |
| Terpenes | ||||
| Penicillium | Azadirachta indica | Azadirachtin | Insecticidal | Kusari, et al. [91] |
| Nodulisporium sp. | Cinnamomum camphora | Camphor | Antimicrobial, topical skin preparations | Suwannarach, et al. [126] |
| Quinones and Xanthones | ||||
| Chaetomium globosum | Hypericum perforatum | Hypericin | Anti-depressant, antimicrobial | Kusari, et al. [91] |
| Gibberella moniliformis | Lawsonia inermis | Lawsone | Cytotoxic | Sarang, et al. [127] |
| Miscellaneous Plant-derived Compounds | ||||
| Aspergillus unguis | Wheat, rye, barley | Azelaic acid | Antimicrobial, anticancer | Kamat, et al. [128] |
| Muscodor vitigenus | Ancistrocladus tectorius | Naphthalene | Antibacterial, insect repellent | Ruangrungsi, et al. [129] |
Table 5: A list of plant-derived compounds from fungal endophytes.
Production of Immunosuppressive Compounds
Immunosuppressive drugs are offered to patients undergoing organ transplantation in order to suppress, minimise, or avoid allograft rejection. Therefore, they are very important in medical management of autoimmune diseases like SLE etc. Because of the rising demand for immunosuppressive treatments, there is a pressing need to accelerate the search for pharmaceuticals that are not only safer but also more trustworthy. This is necessary in order to investigate the issues that are now plaguing the efficacy of these medications. Many investigations have shown that endophytes are capable of producing bioactive chemicals that may suppress the immune system [65, 130]. Chemical analysis is often employed to identify the presence of (-) mycousnine enamine. The endophyte Mycosphaerella sp. was isolated from the china root leaves plant to produce this biomolecule [131]. Cyclosporin A and (-) mycousnine enamine were able to decrease the proliferation of T lymphocytes by reducing the expression of CD25 and CD69 surface activation antigens. This resulted in the inhibition of T cell proliferation [131]. In conjunction to this, nine polyketides were isolated from the mangrove leaves that harbour the endophytic fungus Penicillium sp. ZJ-SY2. These compounds showed promising immunosuppressive properties [130]. Fomitopsisbetulinus, a basidiomycete often linked with wood decay, was the source of Xylarialongipes HFG1018. This fungus was responsible for the production of eighteen new nor-isopimarane diterpenes, which were designated as xylarinorditerpenes A–R (1–18). Some of these compounds had the ability to suppress the immune system [132].
Production of Siderophores
Siderophores are created by many species with the ability to scavenge iron from the environment they are in to render the essential substance easily accessible to the cell. Various species are capable of producing siderophores. To facilitate uptake and utilisation of ferric iron, siderophores are secreted out into the environment to form soluble ferric complexes. Siderophores have very complicated chemical structures, which enables them to create the most powerful iron-chelating complexes. Siderophores are engaged in the process of virulence and play an important role in the mobilisation of iron and other components. Recent research has also shed light on the significant connection between siderophores and the capacity to withstand oxidative stress. Both in agricultural and medical contexts, their applications have been the subject of much research. On the other hand, new research areas are beginning to focus on the use of siderophores as green-iron chelators. Specifically, siderophores are being considered as a potential solution for the protection of cultural assets.
Many microorganisms that live on or around plants, both beneficial and harmful, share a common trait: the synthesis of extracellular siderophores. Siderophores are produced by a number of different strains of fungus. Scots pine and Labrador tea foliar endophytic fungi were studied for their ability to produce antimicrobial and antioxidant compounds via extracellular siderophore production. In vitro production of the siderophore ferricrocin resulted in concentrations ranging from 7.9 to 17.6 micrograms per litre. Ferricrocin was only created by the fungus that also had antibacterial action, and none of the well-known siderophores were found in the broths made by the fungi that produced antioxidants. As a result, the synthesis of ferricrocin is characteristic of certain foliar endophytic fungus, but not all of them. The discovery of ferricrocin in the leaves of Labrador tea lends credence to the hypothesis that this compound may play a function in vivo in the dynamic relationship that exists between the endophyte and the plant host [133].
Conclusion
New and exciting avenues for applied research into plant-microbe interactions can be found by studying the function of fungal endophytes, as these microbes can boost germination rates, enhance seedling health, and increase plant growth. The production of extracellular enzymes, phytohormones, and secondary metabolites can be linked to these abilities. Because of their potential to be used as a biofertilizer to improve plant growth, we should encourage further research into these microbes. An enormous portion of the endophytic population in terrestrial plants is probably still undiscovered. Metagenomic research into plant endophyte populations will reveal new applications and details about these microorganisms, such as the genera, phenotypic traits, and potential roles in seed germination and plant development. There is a need for more study to determine the genetic determinants involved in plant growth promotion and the molecular mechanisms by which plant-endophyte interactions induce defence resistance mechanisms against different types of stresses.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
Grabka R, d’Entremont TW, Adams SJ, Walker AK, Tanney JB, et al. (2022) Fungal Endophytes and Their Role in Agricultural Plant Protection against Pests and Pathogens. Plants (Basel) 11(3): 384.
-
Hassan SED (2017) Plant growth-promoting activities for bacterial and fungal endophytes isolated from medicinal plant of Teucrium polium L. Journal of Advanced Research 8: 687-695.
-
Fontana DC, Paula SD, Torres AG, Souza VHMD, Pascholati SF, et al. (2021) Endophytic Fungi: Biological Control and Induced Resistance to Phytopathogens and Abiotic Stresses. Pathogens 10(5): 570.
-
Wen J, Okyere SK, Wang S, Wang J, Xie L, et al. (2022) Endophytic Fungi: An Effective Alternative Source of Plant-Derived Bioactive Compounds for Pharmacological Studies. J Fungi (Basel) 8(2): 205.
-
Baron NC, Rigobelo EC (2021) Endophytic fungi: a tool for plant growth promotion and sustainable agriculture. Mycology 13(1): 39-55.
-
Chaudhary P, Agri U, Chaudhary A, Kumar A, Kumar G (2022) Endophytes and their potential in biotic stress management and crop production. Front Microbiol 13: 933017.
-
Rashid MdHO, Chung YR (2017) Induction of Systemic Resistance against Insect Herbivores in Plants by Beneficial Soil Microbes. Front Plant Sci 8: 1816.
-
Strobel G, Daisy B (2003) Bioprospecting for Microbial Endophytes and Their Natural Products. Microbiol Mol Biol Rev 67(4): 491-502.
-
Saikkonen K, Wali P, Helander M, Faeth SH (2004) Evolution of endophyte-Plant symbioses. Trends in Plant Science 9(6): 275-280.
-
Hawas UW, El-Beih AA, El-Halawany AM (2012) Bioactive anthraquinones from endophytic fungus Aspergillus versicolor isolated from red sea algae. Arch Pharm Res 35(10): 1749-1756.
-
Samal I, Bhoi TK, Majhi PK, Murmu S, Pradhan AK, et al. (2023) Combatting insects mediated biotic stress through plant associated endophytic entomopathogenic fungi in horticultural crops. Front Plant Sci 13: 1098673.
-
Bamisile BS, Dash CK, Akutse KS, Keppanan R, Wang L (2018) Fungal Endophytes: Beyond Herbivore Management. Front Microbiol 9: 544.
-
Koh S, Hik DS (2007) Herbivory Mediates Grass– Endophyte Relationships. Ecology 88(11): 2752-2757.
-
Zhang J, Lu J, Zhu Y, Shen X, Zhu B, et al. (2024) Roles of endophytic fungi in medicinal plant abiotic stress response and TCM quality development. Chinese Herbal Medicines 16(2): 204-213.
-
Egamberdieva D, Wirth SJ, Alqarawi AA, Allah EFA, Hashem A (2017) Phytohormones and Beneficial Microbes: Essential Components for Plants to Balance Stress and Fitness. Front Microbiol 8: 2104.
-
Yan L, Zhu J, Zhao X, Shi J, Jiang C, et al. (2019) Beneficial effects of endophytic fungi colonization on plants. Appl Microbiol Biotechnol 103(8): 3327-3340.
-
Khan AL, Hussain J, Al-Harrasi A, Al-Rawahi A, Lee IJ (2015) Endophytic fungi: resource for gibberellins and crop abiotic stress resistance. Crit Rev Biotechnol 35(1): 62-74.
-
Khan AL, Hamayun M, Kang SM, Kim YH, Jung HY, et al. (2012) Endophytic fungal association via gibberellins and indole acetic acid can improve plant growth under abiotic stress: an example of Paecilomyces formosus LHL10. BMC Microbiol 12: 3.
-
Jan FG, Hamayun M, Hussain A, Jan G, Iqbal A, et al. (2019) An endophytic isolate of the fungus Yarrowia lipolytica produces metabolites that ameliorate the negative impact of salt stress on the physiology of maize. BMC Microbiol 19(1): 3.
-
Hamayun M, Hussain A, Khan SA, Kim HY, Khan AL, et al. (2017) Gibberellins Producing Endophytic Fungus Porostereum spadiceum AGH786 Rescues Growth of Salt Affected Soybean. Front Microbiol 8: 686.
-
Hamayun MI, Hussain A, Iqbal A, Khan SA, Lee IJ (2020) _Aspergillus niger_ boosted heat stress tolerance in sunflower and soybean via regulating their metabolic and antioxidant system. J Plant Interactions 15(1): 223- 232.
-
Poveda J, Abril-Urias P, Escobar C (2020) Biological Control of Plant-Parasitic Nematodes by Filamentous Fungi Inducers of Resistance: Trichoderma, Mycorrhizal and Endophytic Fungi. Front Microbiol 11: 992.
-
Lugtenberg BJJ, Caradus JR, Johnson LJ (2016) Fungal endophytes for sustainable crop production. FEMS Microbiol Ecol 92(12): fiw194.
-
Zhou W, Wheeler TA, Starr JL, Valencia CU, Sword GA (2018) A fungal endophyte defensive symbiosis affects plant-nematode interactions in cotton. Plant Soil 422: 251-266.
-
Dash CK, Bamisile BS, Keppanan R, Qasim M, Lin Y, et al. (2018) Endophytic entomopathogenic fungi enhance the growth of Phaseolus vulgaris L. (Fabaceae) and negatively affect the development and reproduction of Tetranychus urticae Koch (Acari: Tetranychidae). Microb Pathog 125: 385-392.
-
Wang Y, Mostafa S, Zeng W, Jin B (2021) Function and Mechanism of Jasmonic Acid in Plant Responses to Abiotic and Biotic Stresses. Int J Mol Sci 22(16): 8568.
-
Jaber LR, Enkerli J (2017) Fungal entomopathogens as endophytes: can they promote plant growth? Biocontrol Science and Technology 27(1): 28-41.
-
Jaber LR (2018) Seed inoculation with endophytic fungal entomopathogens promotes plant growth and reduces crown and root rot (CRR) caused by Fusarium culmorum in wheat. Planta 248(6): 1525-1535.
-
Card S, Johnson L, Teasdale S, Caradus J (2016) Deciphering endophyte behaviour: the link between endophyte biology and efficacious biological control agents. FEMS Microbiol Ecol 92(8): fiw 114.
-
Fredenhagen A, Molleyres LP, Böhlendorf B, Laue G (2006) Structure Determination of Neoefrapeptins A to N: Peptides with Insecticidal Activity Produced by the Fungus Geotrichum candidum. J Antibiot (Tokyo) 59(5): 267-280.
-
Krasnoff SB, Keresztes I, Gillilan RE, Szebenyi DME, Donzelli BGG, et al. (2007) Serinocyclins A and B, Cyclic Heptapeptides from _Metarhizium anisopliae_. J Nat Prod 70(12): 1919-1924.
-
Liermann JC, Opatz T, Kolshorn H, Antelo L, Hof C, et al. (2009) Omphalotins E-I, Five Oxidatively Modified Nematicidal Cyclopeptides from _Omphalotus olearius_. Eur J Org Chem 2009(8): 1256-1262.
-
Singh SB, Zink DL, Liesch JM, Dombrowski AW, Darkin-Rattray SJ, et al. (2001) Structure, Histone Deacetylase, and Antiprotozoal Activities of Apicidins B and C, Congeners of Apicidin with Proline and Valine Substitutions. Org Lett 3(18): 2815-2818.
-
Abe M, Imai T, Ishii N, Usui M (2006) Synthesis of Quinolactacide _via_ an Acyl Migration Reaction and Dehydrogenation with Manganese Dioxide, and Its Insecticidal Activities. Biosci Biotechnol Biochem 70(1): 303-306.
-
Ōmura S, Miyadera H, Ui H, Shiomi K, Yamaguchi Y, et al. (2001) An anthelmintic compound, nafuredin, shows selective inhibition of complex I in helminth mitochondria. Proc Natl Acad Sci USA 98(1): 60-62.
-
Kuzma M, Jegorov A, Kacer P, Havlicek V (2001) Sequencing of new beauverolides by high-performance liquid chromatography and mass spectrometry. J Mass Spectrom 36(10): 1108-1115.
-
Bric JM, Bostock RM, Silverstone SE (1991) Rapid in Situ Assay for Indoleacetic Acid Production by Bacteria Immobilized on a Nitrocellulose Membrane. Appl Environ Microbiol 57(2): 535-538.
-
Talukdar R, Tayung K (2019) Antimicrobial Activity of Endophytic Fungi Isolated from Eryngium foetidum, an Ethnomedicinal Plant of Assam. IJPSDR 11(6).
-
Dar RA, Saba I, Shahnawaz M, Sangale MK, Ade AB, et al. (2013) Isolation, purification and characterization of carboxymethyl cellulase (CMCase) from endophytic Fusarium oxysporum producing podophyllotoxin. AER 1(4): 91-96.
-
Szilagyi-Zecchin VJ, Ikeda AC, Hungria M, Adamoski D, Kava-Cordeiro V, et al. (2014) Identification and characterization of endophytic bacteria from corn (Zea mays L.) roots with biotechnological potential in agriculture. AMB Expr 4: 26.
-
Donate-Correa J, León-Barrios M, Pérez-Galdona R (2004) Screening for plant growth-promoting rhizobacteria in Chamaecytisus proliferus (tagasaste), a forage tree-shrub legume endemic to the Canary Islands. Plant and Soil 266(1/2): 261-272.
-
Hankin L, Anagnostakis SL (1975) The Use of Solid Media for Detection of Enzyme Production by Fungi. Mycologia 67(3): 597-607.
-
Schwyn B, Neilands JB (1987) Universal chemical assay for the detection and determination of siderophores. Anal Biochem 160(1): 47-56.
-
Carvalho JOD, Broll V, Martinelli AHS, Lopes FC (2020) Endophytic fungi: positive association with plants. In: Sharma V, et al. (Eds.), Molecular Aspects of Plant Beneficial Microbes in Agriculture. Elsevier, pp: 321-332.
-
Nyoki D, Ndakidemi PA (2014) Effects of Bradyrhizobium japonicum Inoculation and Supplementation with Phosphorus on Macronutrients Uptake in Cowpea (Vigna unguiculata (L.) Walp). AJPS 5(4): 442-451.
-
Rodrı́guez H, Fraga R (1999) Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol Adv 17(4-5): 319-339.
-
Silva AGD (2017) Forms of soil organic phosphorus at black earth sites in the Eastern Amazon. Revista Ciência Agronômica 48(1).
-
Priyadharsini P, Muthukumar T (2017) The root endophytic fungus Curvularia geniculata from Parthenium hysterophorus roots improves plant growth through phosphate solubilization and phytohormone production. Fungal Ecology 27(Part A): 69-77.
-
Wakelin SA, Warren RA, Harvey PR, Ryder MH (2004) Phosphate solubilization by Penicillium spp. closely associated with wheat roots. Biology and Fertility of Soils 40: 36-43.
-
Yadav AN, Sharma D, Gulati S, Singh S, Dey R, et al. (2015) Haloarchaea Endowed with Phosphorus Solubilization Attribute Implicated in Phosphorus Cycle. Sci Rep 5: 12293.
-
Rinu K, Sati P, Pandey A (2014) _Trichoderma gamsii_ (NFCCI 2177): A newly isolated endophytic, psychrotolerant, plant growth promoting, and antagonistic fungal strain. J Basic Microbiol 54(5): 408-417.
-
Nath R, Sharma GD, Barooah M (2015) Plant Growth Promoting Endophytic Fungi Isolated From Tea (Camellia Sinensis) Shrubs Of Assam, India. AEER 13(3): 877-891.
-
Khan SA, Hamayun M, Yoon H, Kim HY, Suh SJ, et al. (2008) Plant growth promotion and Penicillium citrinum. BMC Microbiol 8: 231.
-
You YH, Kwak TW, Kang SM, Lee MC, Kim JG (2015) _Aspergillus clavatus_ Y2H0002 as a New Endophytic Fungal Strain Producing Gibberellins Isolated from _Nymphoides peltata_ in Fresh Water. Mycobiology 43(1): 87-91.
-
Khan AR, Ullah I, Waqas M, Shahzad R, Hong SJ, et al. (2015) Plant growth-promoting potential of endophytic fungi isolated from Solanum nigrum leaves. World J Microbiol Biotechnol 31(9): 1461-1466.
-
Carroll G (1988) Fungal Endophytes in Stems and Leaves: From Latent Pathogen to Mutualistic Symbiont. Ecology 69(1): 2-9.
-
Yadav C (2018) New and Future Developments in Microbial Biotechnology and Bioengineering: penicillium system properties and applications. Elsevier, Amsterdam.
-
Benjamin S, Pandey A (1998) Candida rugosa lipases: Molecular biology and versatility in biotechnology. Yeast 14(12): 1069-1087.
-
Rosa GDI, Peralta-Videa JR, Montes M, Parsons JG, Cano-Aguilera I, et al. (2004) Cadmium uptake and translocation in tumbleweed (Salsola kali), a potential Cd-hyperaccumulator desert plant species: ICP/OES and XAS studies. Chemosphere 55(9): 1159-1168.
-
Soleimani M, Afyuni M, Hajabbasi MA, Nourbakhsh F, Sabzalian MR, et al. (2010) Phytoremediation of an aged petroleum contaminated soil using endophyte infected and non-infected grasses. Chemosphere 81(9): 1084- 1090.
-
Zhang W, Niu Y, Li YX, Zhang F, Zeng RJ (2020) Enrichment of hydrogen-oxidizing bacteria with nitrate recovery as biofertilizers in the mixed culture. Bioresour Technol 313: 123645.
-
Ma Y, Rajkumar M, Luo YM, Freitas H (2011) Inoculation of endophytic bacteria on host and non-host plants— Effects on plant growth and Ni uptake. J Hazard Mater 195: 230-237.
-
Martin JF, McDaniel LE (1977) Production of Polyene Macrolide Antibiotics. Adv Appl Microbiol 21: 1-52.
-
Kirkpatrick P, Raja A, LaBonte J, Lebbos J (2003) Daptomycin. Nat Rev Drug Discov 2(12): 943-944.
-
Wang WX, Cheng GG, Li ZH, Ai HL, He J, et al. (2019) Curtachalasins, immunosuppressive agents from the endophytic fungus _Xylaria_ cf. _curta_. Org Biomol Chem 17(34): 7985-7994.
-
Strobel GA, Miller RV, Martinez-Miller C, Condron MM, Teplow DB, et al. (1999) Cryptocandin, a potent antimycotic from the endophytic fungus Cryptosporiopsis cf. quercina. Microbiology (Reading) 145(Pt 8): 1919-1926.
-
Li JY, Harper JK, Grant DM, Tombe BO, Bashyal B, et al. (2001) Ambuic acid, a highly functionalized cyclohexenone with antifungal activity from Pestalotiopsis spp. and Monochaetia sp. Phytochemistry 56(5): 463-468.
-
Ballio A, Barra D, Bossa F, Collina A, Grgurina I, et al. (1991) Syringopeptins, new phytotoxic lipodepsipeptides of _Pseudomonas syringae_ pv. _syringae_. FEBS Lett 291(1): 109-112.
-
Harrison L, Teplow DB, Rinaldi M, Strobel G (1991) Pseudomycins, a family of novel peptides from Pseudomonas syringae possessing broad-spectrum antifungal activity. J Gen Microbiol 137(12): 2857-2865.
-
Harper JK, Mulgrew AE, Li JY, Barich DH, Strobel GA, et al. (2001) Characterization of Stereochemistry and Molecular Conformation Using Solid-State NMR Tensors. J Am Chem Soc 123(40): 9837-9842.
-
Lee JC, Yang X, Schwartz M, Strobel G, Clardy J (1995) The relationship between an endangered North American tree and an endophytic fungus. Chem Biol 2(11): 721- 727.
-
Strobel GA, Dirkse E, Sears J, Markworth C (2001) Volatile antimicrobials from Muscodor albus, a novel endophytic fungus. Microbiology (Reading) 147(Pt 11): 2943-2950.
-
Brady SF, Wagenaar MM, Singh MP, Janso JE, Clardy J (2000) The Cytosporones, New Octaketide Antibiotics Isolated from an Endophytic Fungus. Org Lett 2(25): 4043-4046.
-
Pongcharoen W, Rukachaisirikul V, Phongpaichit S, Kühn T, Pelzing M, et al. (2008) Metabolites from the endophytic fungus Xylaria sp. PSU-D14. Phytochemistry 69(9): 1900-1902.
-
Pittayakhajonwut P, Suvannakad R, Thienhirun S, Prabpai S, Kongsaeree P, et al. (2005) An anti-herpes simplex virus-type 1 agent from Xylaria mellisii (BCC 1005). Tetrahedron Letters 46(8): 1341-1344.
-
Li E, Jiang L, Guo L, Zhang H, Che Y (2008) Pestalachlorides A-C, antifungal metabolites from the plant endophytic fungus Pestalotiopsis adusta. Bioorg Med Chem 16(17): 7894-7899.
-
Buatong J, Phongpaichit S, Rukachaisirikul V, Sakayaroj J (2011) Antimicrobial activity of crude extracts from mangrove fungal endophytes. World J Microbiol Biotechnol 27: 3005-3008.
-
Xu L, Zhou L, Zhao J, Li J, Li X, et al. (2008) Fungal endophytes from Dioscorea zingiberensis rhizomes and their antibacterial activity. Lett Appl Microbiol 46(1): 68-72.
-
Mao Z, Zhang W, Wu C, Feng H, Peng Y, et al. (2021) Diversity and antibacterial activity of fungal endophytes from Eucalyptus exserta. BMC Microbiol 21(1): 155.
-
Cragg GM, Grothaus PG, Newman DJ (2009) Impact of Natural Products on Developing New Anti-Cancer Agents. Chem Rev 109(7): 3012-3043.
-
Joseph B, Priya RM (2011) Bioactive Compounds from Endophytes and their Potential in Pharmaceutical Effect: A Review. American J of Biochemistry and Molecular Biology 1(3): 291-309.
-
Alvin A, Miller KI, Neilan BA (2014) Exploring the potential of endophytes from medicinal plants as sources of antimycobacterial compounds. Microbiol Res 169(7- 8): 483-495.
-
Xie S, Ogden A, Aneja R, Zhou J (2016) Microtubule- Binding Proteins as Promising Biomarkers of Paclitaxel Sensitivity in Cancer Chemotherapy. Med Res Rev 36(2): 300-312.
-
Kharwar RN, Mishra A, Gond SK, Stierle A, Stierle D (2011) Anticancer compounds derived from fungal endophytes: their importance and future challenges. Nat Prod Rep 28(7): 1208-1228.
-
Ran X, Zhang G, Li S, Wang J (2017) Characterization and antitumor activity of camptothecin from endophytic fungus _Fusarium solani_ isolated from _Camptotheca_ _acuminate_. Afr Health Sci 17(2): 566-574.
-
Giri A, Narasu ML (2000) Production of podophyllotoxin from Podophyllum hexandrum: a potential natural product for clinically useful anticancer drugs. Cytotechnology 34(1-2): 17-26.
-
Jiao RH, Xu S, Liu JY, Ge HM, Ding H, et al. (2006) Chaetominine, a Cytotoxic Alkaloid Produced by Endophytic _Chaetomium_ sp. IFB-E015. Org Lett 8(25): 5709-5712.
-
Sappapan R, Sommit D, Ngamrojanavanich N, Pengpreecha S, Wiyakrutta S, et al. (2008) 11-Hydroxymonocerin from the Plant Endophytic Fungus _Exserohilum rostratum_. J Nat Prod 71(9): 1657-1659.
-
Zuccaro A, Lahrmann U, Langen G (2014) Broad compatibility in fungal root symbioses. Curr Opin Plant Biol 20: 135-145.
-
Tan RX, Zou WX (2001) Endophytes: a rich source of functional metabolites. Nat Prod Rep 18(4): 448-459.
-
Kusari S, Lamshöft M, Zühlke S, Spiteller M (2008) An Endophytic Fungus from _Hypericum perforatum_ that Produces Hypericin. J Nat Prod 71(2): 159-162.
-
Huang Z, Guo Z, Yang R, Yin X, Li X, et al. (2009) Chemistry and cytotoxic activities of polyketides produced by the mangrove endophytic fungus Phomopsis SP. ZSU-H76. Chem Nat Compd 45: 625-628.
-
Isaka M, Jaturapat A, Rukseree K, Danwisetkanjana K, Tanticharoen M, et al. (2001) Phomoxanthones A and B, Novel Xanthone Dimers from the Endophytic Fungus _Phomopsis_ Species. J Nat Prod 64(8): 1015-1018.
-
Guo B, Wang Y, Sun X, Tang K (2008) Bioactive natural products from endophytes: a review. Prikl Biokhim Mikrobiol 44(2): 153-158.
-
Kralj A, Kehraus S, Krick A, Eguereva E, Kelter G, et al. (2006) Arugosins G and H: Prenylated Polyketides from the Marine-Derived Fungus _Emericella_ _n_ _idulans_ var. _a_ _cristata_. J Nat Prod 69(7): 995-1000.
-
Wani MC, Taylor HL, Wall ME, Coggon P, McPhail AT (1971) Plant antitumor agents. VI. Isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia. J Am Chem Soc 93(9): 2325- 2327.
-
Logan DC (2007) The mitochondrial compartment. J Exp Bot 58(1): 1225-1243.
-
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48(12): 909-930.
-
Foyer CH, Noctor G (2000) Tansley Review No. 112: Oxygen processing in photosynthesis: regulation and signalling. New Phytologist 146(3): 359-388.
-
Apel K, Hirt H (2004) Reactive Oxygen Species: Metabolism, Oxidative Stress, and Signal Transduction. Annu Rev Plant Biol 55: 373-399.
-
Jaspers P, Kangasjärvi J (2010) Reactive oxygen species in abiotic stress signaling. Physiol Plant 138(4): 405-413.
-
Overmyer K, Brosché M, Kangasjärvi J (2003) Reactive oxygen species and hormonal control of cell death. Trends Plant Sci 8(7): 335-342.
-
Gara LD, Locato V, Dipierro S, Pinto MCD (2010) Redox homeostasis in plants. The challenge of living with endogenous oxygen production. Respir Physiol Neurobiol 173: S13-S19.
-
Foyer CH, Noctor G (2011) Ascorbate and Glutathione: The Heart of the Redox Hub. Plant Physiol 155(1): 2-18.
-
Paul S, Bhagobaty RK, Nihalani MC, Joshi SR (2020) Characterization of oleaginous endophytic fungi of biodiesel plants as potential biofuel minifactories. Biomass and Bioenergy 142: 105750.
-
Shen B (2015) A New Golden Age of Natural Products Drug Discovery. Cell 163(6): 1297-1300.
-
Newsham KK (2011) A meta‐analysis of plant responses to dark septate root endophytes. New Phytol 190(3): 783-793.
-
Calixto JB (2019) The role of natural products in modern drug discovery. An Acad Bras Ciênc 91(Suppl 3): e20190105.
-
Wilson BAP, Thornburg CC, Henrich CJ, Grkovic T, O’Keefe BR (2020) Creating and screening natural product libraries. Nat Prod Rep 37(7): 893-918.
-
Strobel G (2018) The Emergence of Endophytic Microbes and Their Biological Promise. J Fungi (Basel) 4(2): 57.
-
Abdel-Razek AS, El-Naggar ME, Allam A, Morsy OM, Othman SI (2020) Microbial Natural Products in Drug Discovery. Processes 8(4): 470.
-
Wu H, Yang HY, You XL, Li YH (2013) Diversity of endophytic fungi from roots of Panax ginseng and their saponin yield capacities. SpringerPlus 2(1): 107.
-
Yang K, Liang J, Li Q, Kong X, Chen R, et al. (2013) Cladosporium cladosporioides XJ-AC03, an aconitine- producing endophytic fungus isolated from Aconitum leucostomum. World J Microbiol Biotechnol 29(5): 933- 938.
-
Devari S, Jaglan S, Kumar M, Deshidi R, Guru S, et al. (2014) Capsaicin production by Alternaria alternata, an endophytic fungus from Capsicum annum; LC–ESI–MS/ MS analysis. Phytochemistry 98: 183-189.
-
Kumara PM, Soujanya KN, Ravikanth G, Vasudeva R, Ganeshaiah KN, et al. (2014) Rohitukine, a chromone alkaloid and a precursor of flavopiridol, is produced by endophytic fungi isolated from Dysoxylum binectariferum Hook.f and Amoora rohituka (Roxb). Wight & Arn. Phytomedicine 21(4): 541-546.
-
Zaher AM, Moharram AM, Davis R, Panizzi P, Makboul MA, et al. (2015) Characterisation of the metabolites of an antibacterial endophyte _Botryodiplodia theobromae_ Pat. of _Dracaena draco_ L. by LC–MS/MS. Nat Prod Res 29(24): 2275-2281.
-
Yamazaki T, Tokiwa T (2010) Isofraxidin, a Coumarin Component from Acanthopanax senticosus, Inhibits Matrix Metalloproteinase-7 Expression and Cell Invasion of Human Hepatoma Cells. Bio Pharm Bull 33(10): 1716- 1722.
-
Cheng C, Li W, Lin M, Yang ST (2019) Metabolic engineering of Clostridium carboxidivorans for enhanced ethanol and butanol production from syngas and glucose. Bioresour Technol 284: 415-423.
-
Oliveira CM, Regasini LO, Silva GH, Pfenning LH, Young MCM, et al. (2011) Dihydroisocoumarins produced by Xylaria sp. and Penicillium sp., endophytic fungi associated with Piper aduncum and Alibertia macrophylla. Phytochemistry Letters 4(2): 93-96.
-
Shukla S, Gupta S (2010) Apigenin: A Promising Molecule for Cancer Prevention. Pharm Res 27(6): 962- 978.
-
Zhao Y, Zhao T, Jia H, Li X, Zhu Z, et al. (2017) Optimization of the composition of mixed entrainer for economic extractive distillation process in view of the separation of tetrahydrofuran/ethanol/water ternary azeotrope. Optimization of the composition of mixed entrainer. J Chem Technol Biotechnol 92(9): 2433-2444.
-
Wang X (2020) Groundless Research Published on the International Journal of Plant Sciences. VP 6(4): 167- 169.
-
Chapela IH, Petrini O, Hagmann L (1991) Monolignol glucosides as specific recognition messengers in fungus- plant symbioses. Physiological and Molecular Plant Pathology 39(4): 289-298.
-
Kong P, Zhang L, Guo Y, Lu Y, Lin D (2014) Phillyrin, a Natural Lignan, Attenuates Tumor Necrosis Factor α-Mediated Insulin Resistance and Lipolytic Acceleration in 3T3-L1 Adipocytes. Planta Med 80(11): 880-886.
-
Parthasarathy R, Sathiyabama M (2014) Gymnemagenin-producing Endophytic Fungus Isolated from a Medicinal Plant Gymnema sylvestre R.Br. Appl Biochem Biotechnol 172(6): 3141-3152.
-
Suwannarach N, Kumla J, Bussaban B, Nuangmek W, Matsui K, et al. (2013) Biofumigation with the endophytic fungus Nodulisporium spp. CMU-UPE34 to control postharvest decay of citrus fruit. Crop Protection 45: 63-70.
-
Sarang H, Rajani P, Vasanthakumari MM, Kumara PM, Siva R, et al. (2017) An endophytic fungus, Gibberella moniliformis from Lawsonia inermis L. produces lawsone, an orange-red pigment. Antonie van Leeuwenhoek 110(7): 853-862.
-
Kamat S, Kumari M, Taritla S, Jayabaskaran C (2020) Endophytic Fungi of Marine Alga From Konkan Coast, India—A Rich Source of Bioactive Material. Front Mar Sci 7: 31.
-
Ruangrungsi N, Wongpanich V, Tantivatana P, Cowe HJ, Cox PJ, et al. (1985) Traditional Medicinal Plants of Thailand, V. Ancistrotectorine, a New Naphthalene- Isoquinoline Alkaloid from Ancistrocladus tectorius. J Nat Prod 48(4): 529-535.
-
Liu H, Chen S, Liu W, Liu Y, Huang X, et al. (2016) Polyketides with Immunosuppressive Activities from Mangrove Endophytic Fungus Penicillium sp. ZJ-SY2. Mar Drugs 14(12): 217.
-
Wang LW, Wang JL, Chen J, Chen JJ, Shen JW, et al. (2017) A Novel Derivative of (-)mycousnine Produced by the Endophytic Fungus Mycosphaerella nawae, Exhibits High and Selective Immunosuppressive Activity on T Cells. Front Microbiol 8: 1251.
-
Chen HP, Zhao ZZ, Cheng GG, Zhao K, Han KY, et al. (2020) Immunosuppressive Nor-isopimarane Diterpenes from Cultures of the Fungicolous Fungus _Xylaria longipes_ HFG1018. J Nat Prod 83(2): 401-412.
-
Kajula M, Tejesvi MV, Kolehmainen S, Mäkinen A, Hokkanen J, et al. (2010) The siderophore ferricrocin produced by specific foliar endophytic fungi in vitro. Fungal Biol 114(2-3): 248-254.
- Antifungal Activity of New Acetophenone Derivatives
- Interconnected Microbiomes Human Health Within an Environmental Framework
- Silkworm-Based Vaccine Production for H5N1: A One Health Approach to Pandemic Preparedness
- Microbial Diversity and Lipolytic Activity of Bacteria and Fungi from Oil-Contaminated Sites in Makurdi Metroplois
- Antibiotic Resistance Profile of Bacteria Isolated at the Central Laboratory of the National Hospital Center of Nouakchott
- Epidemiology and Sensitivity to Antibiotics of Germs Isolated from Blood Cultures in the Laboratory of the National Hospital Center of Nouakchott-Mauritania