Influence of Inoculum Concentration on In Vivo Incubation Period of Emmia lacerata, Pathogenesis and Management of Wilt in Pepper (Capsicum annuum L.)
Emerging infectious disease, caused by Emmia lacerata (a host-jumping phytopathogen with medical significance), is a potential threat to production and consumption of pepper. Characterizing the pathogenesis of E. lacerata and management of Emmia Wilt (EW) in pepper is imperative. In vitro antifungal property of Bacillus velezensis EBs02 and B. thuringiensis EBs04 against E. lacerata FMIB29 was screened on dual-culture agar. Effect of inoculum dose on incubation period, severity of EW and its biocontrol was investigated. Bacillus velezensis EBs02 expressed the highest mycelial control (68.33%) against FMIB29. A 2-week incubation period was observed for E. lacerata in pepper and plants infected with 108 spore/mL exhibited the highest EW severity index (45.01%). Highest in vivo disease control (72.75%) was observed in pepper treated with EBs02 and differences in inoculum dose of FMIB29 neither influenced wilt severity nor the biocontrol potential of Bacillus strains. However, both Bacillus strains drastically reduced the manifestation of wilt in pepper.
Adedire OM¹, Adedire DE ², Owoeye TE³ and Aina A⁴
¹School of Agriculture, Federal College of Agriculture, Nigeria
Introduction
Pepper (Capsicum species) fruit is an outstanding source of micro and macronutrients for several households all over the world [1, 2]. It is a reservoir of bioactive metabolites, such as carotenoids, phenolics, flavonoids, as well as iron, dietary source of provitamin A, ascorbic acid (vitamin C), and other antioxidants [3]. These metabolites have been reported to improve cardiovascular ailment and possibly, prostate cancer; while carotenoids in pepper produce its characteristic aroma and flavor [2]. This makes pepper a vegetable of interest for farmers and a spice of choice for cuisines, especially, in southwestern States of Nigeria.
Unfortunately, phytopathogens militate against the production of fruits and vegetables globally [4]; consequently, frustrating the concept of global food security and nutrition. If poorly managed, pathogens could cause up to 80 - 100% of pepper loss on the field, while 20 –25% of harvested fruits are damaged by rot-causing microbes [5]. Nevertheless, commercial farmers are recording significant progress in the management of common pathogens in pepper. However, emerging infectious diseases of pepper constitute an impending threat to the production of this economically significant crop.
In their study on the pathogenicity and host range of Alternaria alternata, Balamurugan and Kumar described the intensity and aggressiveness of the pathogen on solanaceous crops. Xhemali B, et al. [6] also documented the first report on Colletotrichum scovillei causing anthracnose in pepper fruits. Typical symptoms of C. scovillei-associated anthracnose were observed in Capsicum annum cv. Somborka cultivated in different fields of Peja and Rahovec, Kosovo. In 2020 Emmia lacerata was isolated from a tomato farm along Ile-tuntun/ Idi-ishin area, Ibadan, Oyo State, Nigeria (7°24′N; 3°50′E) [7]. The first report on the pathogenicity (chlorosis, necrosis and wilt) of E. lacerata in tomato was thereafter documented [8]. Emmia lacerata equally infected pepper plants cultivated at the Horticultural Unit of the Federal College of Agriculture, Ibadan (7°22ʹN; 3°50ʹE), both in 2021 and 2022.
In addition to their saprophytic potential, Emmia lacerata has been reported as an endophyte and respiratory parasite [9]. In their report on the biostimulation of Michelia macclurei, Pan X, et al. [10] reported a clone-specific improvement in nutrient uptake and a consequential growth promotion caused by E. lacerata SR5. This strain was thereafter proposed as a potential biofertilizer. Through clinical evidence and molecular sequencing, E. lacerata was identified as an emerging invasive mold, responsible for fungal pneumonia and allergies, especially in immunocompromised patients in Korea [9]. Infected patients were presented with occasional manifestation of hemoptysis. Emmia lacerata has been associated with lethal wilt, as well as root and stem rot of olive trees, originally caused by Dematophora necatrix in Central Italy [11]. Although, Emmia lacerata has been investigated as a soil-borne mycopathogen of tomato plant [8], their virulence and management in other solanaceous crops like pepper and eggplants have not been investigated.
Antimycotic Bacillus strains (EBs02 and EBs04) were collected from the Microbiology Unit, Federal College of Agriculture, and Ibadan, Nigeria. They were identified as Bacillus velezensis and Bacillus thuringiensis (Figure 1) and assigned genbank numbers OP975754 and OP975750, respectively. These strains have been reported to express in vitro and in vivo inhibitory potentials against Fusarium solani in maize [13]. Emmia species, originally isolated from wilting tomato plants [7], were acquired from the Seed Health Laboratory of National Center for Genetic Resources and Biotechnology (SHL-NACGRAB), Ibadan, Nigeria. The fungal strain was identified as E. lacerata (Figure 2), with an assigned genbank number MZ959067. Bacillus species were maintained on Nutrient Agar (NA), while E. lacerata was cultivated on Potato Dextrose Agar (PDA).
However, emerging infectious diseases could be very difficult to control. Conventional control practices, deployed without adequate characterization of novel pathogens, may not only fail but also predispose host crops to more devastating microbial attack [12]. Currently, there is a dearth of requisite information on the pathogenesis of Emmia wilt in Pepper, the pattern of pathogen (E. lacerata) transmission and influence of inoculum concentration on management strategies. This research work was designed to establish the pathogenesis of E. lacerata and determine the effect of inoculum dose on the incubation period and management of Emmia wilt in pepper plant.
Materials and Methods
Experimental Sites and Sources of Microbial Strains
The in vitro and screenhouse experiments were carried out between February - April, 2023 and July – September, 2023, respectively. The experiments were conducted at a Research-Extension Unit of the Federal College of Agriculture, Ibadan (7°25’N; 3°50’E). Molecular identification of microbial strains was done at Bioscience Laboratory of the International Institute of Tropical Agriculture, Ibadan.
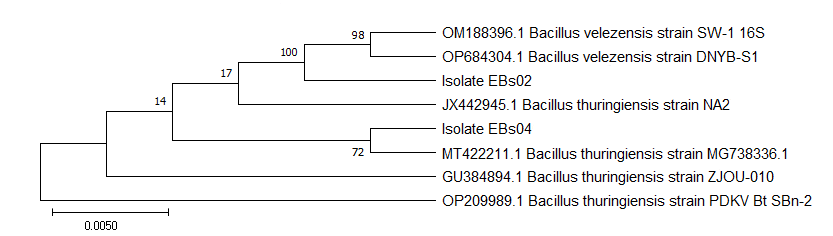
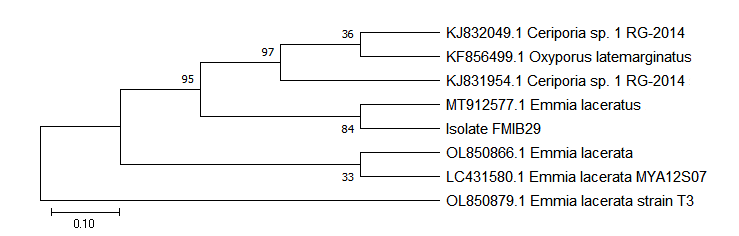
Effect of Inoculum Dose on Pathogenicity and Incubation of Emmia lacerata in Pepper
This was conducted to investigate the influence of spore concentration on disease establishment and severity of wilt in pepper. Such information will be necessary for soil diagnosis before cultivation, as well as subsequent investigation on the impact of soil fungal population on accumulation of E. lacerata, especially, in asymptomatic hosts with medical significance.
Sandy-loam soil samples were collected from the Bora Farm of Federal College of Agriculture, Moor Plantation, Ibadan (7°22’32.0”N; 3°50’49.7”E) and subsequently steam- sterilised. Seeds of C. annuum (acquired from the Seed Store of the Institute of Agricultural Research and Training, Ibadan) were surface-sterilised with 2% NaOCl, and sown in 7 kg steam-sterilised potted soil. Emmia lacerata was cultivated in Potato Dextrose Broth (PDB) for 3 days at 30°C to generate spores. The rhizosphere of 10-day-old pepper seedlings was inoculated with 1 mL of different inoculum doses (106, 107 and 108 spores/mL) of the pathogen [14]. Infected seedlings were observed for 6 weeks after sowing to determine the period between inoculation and manifestation of wilt symptoms (in vivo incubation duration). Disease severity rating and percentage severity index were determined using the severity scale described by Bindal S, et al. [15] as 0 = healthy plant to 5 = dead plant stump.
R Ip Wilt severity index 100 T Sh × = × × ∑
Where R = severity score; Ip = number of infected plants; T = total number of plants under consideration, and Sh = highest severity score (5).
In vitro Inhibitory Assay of Bacillus Strains Against Emmia lacerata
Characterization of antifungal efficacy of biocontrol strains against mycopathogens is an essential step to select inhibitory Bacillus species for subsequent in vivo studies.
The antimycotic property of selected Bacillus strains against E. lacerata was determined through a dual-culture confrontation assay, as described by Cao Y, et al. [16]. The growing edge (5 mm) of 5-day-old E. lacerata was cut using a cork-borer and inoculated at the centre of PDA in a Petri dish. After 24 hours of incubation at 30°C, a day-old strain of each Bacillus species was spot inoculated on the plate, 3.5 cm away from the centre (containing the E. lacerata) [16]. Positive control plates (without Bacillus) and plates with dual cultures were incubated for 6 days at 30°C. Mycelial control was determined using the formula described by Hammami R, et al. [17]:
( ) MD Mc
Mycelial inhibition %
$$ ) = \frac {\mathrm {M D} - \mathrm {M c}}{\mathrm {M D}} \times 1 0 0 $$ Where MD = mycelial diameter on control plate and Mc = mycelial diameter on dual culture plate.
Effect of Inoculum Dose on the Biocontrol of Emmia lacerata with Bacillus Strains
One of the factors affecting the adoption of commercially available biocontrol materials in crop production is the lack of graduation, targeted towards specific pathogens. The efficacy of whole-cell biocontrol strains could be influenced by population of soil pathogens. Hence, depending on the outcome of initial soil diagnosis, a dose-dependent management procedure may be necessary. To determine the effect of inoculum dose on Emmia wilt management.
The rhizosphere of 10-day-old pepper seedlings was inoculated with 1 mL of different inoculum doses (106, 107 and 108 spores/mL) of the pathogen [14], as earlier described. Twenty-four hours after seedling inoculation with the pathogen, rhizosphere of hosts was treated with each Bacillus strain (108 cfu/mL). The experiment was arranged in a completely randomized design containing 3 concentrations of inoculum and two Bacillus strains. Positive control plants were uninoculated and negative (infected) control plants were treated with water. The treatments were replicated 4 times and the influence of inoculum size on wilt-biocontrol potential of selected Bacillus strains was determined 4 weeks after plant inoculation (6 weeks after sowing). Disease severity was established as illustrated by Bindal S, et al. [15], while percentage wilt control (PWC) was determined as described below.
DSC DST Wilt control 100 DSC − = ×
Where DSC = wilt severity index of negative control plant and DST = wilt severity index of Bacillus-treated plant [18].
Statistical Analysis
Data were analysed using both descriptive and inferential analysis. Replicates were subjected to analysis of variance, while means were separated with Duncan’s Multiple Range Test at 5% level of significance, with the statistical package for social sciences (version 25.0).
Results and Discussion
Effect of Inoculum Concentration on Pathogenicity and Incubation Duration of Emmia lacerata in Pepper
Emmia lacerata FMIB29 caused wilt disease, through rhizosphere inoculation, in pepper plant. Manifestation of leaf chlorosis (Figure 3) was observed in infected seedlings 2 weeks post-inoculation with different concentrations of E. lacerata. The period between plant inoculation and the expression of wilt symptoms (2 weeks) was regarded as the incubation period for the pathogen in infected pepper seedlings. Chlorotic plants gradually became necrotic, and this resulted in crown wilt. Differences in inoculum dose, as used in this experiment, neither affected the incubation period observed for E. lacerata in pepper nor the severity of the disease (Table 1). Although, at 6 after sowing, plants inoculated with 108 spore/mL expressed the highest wilt disease severity index (45.01%), this was not significantly higher than the severity observed in plant sets inoculated with lower pathogen concentrations (107 and 106 spore/ mL). Control plants, treated with sterile distilled water, in place of Emmia species, expressed no wilt symptom through the course of the test.
As previously observed in tomato plants [8], the phytopathogenicity of E. lacerata was demonstrated (in pepper) in this study. However, the medical significance of E. lacerata, a fungus previously reported as wood-decaying organism, was established by Lee J, et al. [9]. The organism was described as an emerging human pathogen associated with fungal pneumonia. Supportive clinical pathogenicity assessment included chest consolidation, nodular opacity, air-fluid level, pleural effusions and cavitation. In contrast, the symbiotic potential of E. lacerata strains (as biocontrol agents) was investigated against multiple phytopathogenic fungi and oomycetes [19]. Emmia lacerata HG2011 was reported to successfully control eggplant stem-blight and cucumber vine-blight in infected plants. The occurrence of different physiological forms of E. lacerata might be as a result of horizontal gene transfer between pathogenic and non-pathogenic strains of the species [20].
Mobile pathogenicity chromosomes can be transferred between different morphological forms of the same or different microbial species. A typical example of interspecific virulence gene transfer was demonstrated between Stagonospora nodorum and Pyrenophora triti-ci- repentis [21]. However, occurrence of intraspecific, mobile pathogenicity-chromosome transfer was established between non-pathogenic and virulent strains of Fusarium species [22]. Such transfer of virulence conferred on the non-pathogenic strains the ability to infect tomato host. Consequently, the awareness of physiological forms with different host specificity is imperative in the application of E. lacerata as endosymbiont or as biocontrol agent against phytopathogens. This would guide against the possibility of cross-infection (especially, in immunocompromised human hosts) caused by the consumption of fruits produced by infected or inoculated plants.
Concentration of Emmia inoculum appeared not to significantly affect the incubation period and wilt severity in pepper plant; this could be as a result of inoculum threshold of the pathogen. Chatterton S, et al. [14] investigated the inoculum dose-disease response relationship for pea root- rot pathogen and reported the influence of soil type and microbial interaction on the severity of rot, especially, beyond the inoculum threshold of Aphanomyces euteiches in pea. Increase in rhizosphere concentration of phytopathogens (beyond inoculum threshold) could initiate chemical signals, leading to the expression or activation of phytoalexins against the invading pathogen [23]. This will militate against further increment in wilt severity, as observed for 107 and 108 spore/ mL E. lacerata in pepper. Although, such expression might not inhibit pathogen penetration and the consequential occurrence of wilt, it would interfere with the severity of disease expressed by the infected host.
In vitro Mycelial Inhibition of Emmia lacerata by Bacillus Strains
At 7 days after inoculation on PDA, E. lacerata FMIB29 (control) had a mycelial diameter of 66.00 mm (Table 2).
Bacillus velezensis EBs02 expressed the highest percentage mycelial control (68.33) against E. lacerata FMIB29; however, this was not significantly different from the average mycelial control (39.56%) observed for Bacillus thuringiensis EBs04. Antimycotic Bacillus species have potential to enhance crop productivity. Conventionally, Bacillus strains are screened for their ability to inhibit potential phytopathogens, usually, through the production of metabolites.

| Concentration of Emmia species | Disease severity index (%) | |||
|---|---|---|---|---|
| 1WAI | 2WAI | 3WAI | 2WAI | |
| 106 spore/mL | 0±0.00a | 15.00±5.77ab | 21.66±1.67a | 26.66±4.41a |
| 107 spore/mL | 0±0.00a | 22.33±6.74a | 26.66±9.28a | 33.33±8.82a |
| 108 spore/mL | 0±0.00a | 22.33±6.74a | 28.33±10.14a | 45.01±5.77a |
| Control | 0±0.00a | 0±0.00c | 0±0.00b | 0±0.00b |
Table 1: ** Pathogenicity of Emmia lacerata in pepper.
WAI: Weeks after inoculation Mean values (± standard error). Means followed by the same letter(s) within a column are not significantly different at 5% probability level according to Duncan’s Multiple Range Test. Table 1: Pathogenicity of Emmia lacerata in pepper.
These extracellular bioactive components could serve as safer alternatives to chemical pesticides. Jan F, et al. [24] investigated the in vitro properties of Bacillus subtilis FJ3 and established the biocontrol and plant growth promoting potential of the strain. Mycelial inhibition potential of B. subtilis FJ3 was attributed to the production of hydrolytic enzymes, iturin, fengycin, biofilm and surfactin. In a similar study on the inhibitory potentials of B. velezensis, Sicuia OA, et al. [25] reported a broad spectrum of antifungal activity against phytopathogens such as Fusarium oxysporum, F. solani, F. equiseti, F. proliferatum, Penicillium expansum and strains of Aspergillus species. The antifungal potential of B. velezensis was unaffected by the features of in vitro screening (whole-cell, cell-free or volatile organic compound assay). Bacillus thuringiensis CHGP12 was also reported to produce in vitro antifungal lipopeptides and exhibited a high inhibitory potential (3.45 cm) against the mycelial growth of wilt pathogen in chickpea [26]. In addition to its in vitro properties, B. thuringiensis CHGP12 also induced a 40% reduction in wilt severity and improved the performance of infected chickpeas.
| Mycelial growth (mm) (7DAI) | Mycelial inhibition (7DAI) | Control (%) (7DAI) | |
|---|---|---|---|
| B. velezensis EBs02 | 20.33±0.33c | 0.68±0.02a | 68.33±1.67a |
| B. thuringiensis EBs04 | 46.33±1.86b | 0.39±0.17a | 39.56±16.13a |
| Control | 66.00±0.58a | 0.00±0.00b | 0.00±0.00b |
Table 2: ** In vitro mycelial inhibition of Emmia lacerata by Bacillus strains.
DAI: Days after inoculation Mean values (± standard error). Means followed by the same letter(s) within a column are not significantly different at 5% probability level according to Duncan’s Multiple Range Test. Table 2: In vitro mycelial inhibition of Emmia lacerata by Bacillus strains.
Effect of Bacillus Treatment on the Severity of Pepper Wilt
Bacillus strains (B. velezensis EBs02, B. thuringiensis EBs04), used as a single treatment or in combined application, significantly reduced wilt severity in infected pepper plant (Table 3). Lowest wilt severity (0.75) was observed in Emmia-inoculated plants (at 106 cfu/mL) treated with B. velezensis EBs02; however, this was not significantly higher than other inoculum treatments. Consequently, both Bacillus strains have not potentiated the wilt-biocontrol activities of one another. At the inoculum dose of 108 cfu/ mL, Bacillus EBs02 also expressed the highest wilt control, with a wilt severity score of 1.13; however, this was not significantly different from the severity scores observed for other treatments. Similar to the effect of inoculum dose on pathogenicity of E. lacerata in pepper plant, concentrations of the pathogen did not significantly influence the biocontrol potential of selected Bacillus strains. Highest wilt control (72.75%, 54.50% and 64.00%) determined for Bacillus EBs02, Bacillus EBs04 and the combination of both strains, respectively was expressed in pepper plants inoculated with the lowest concentration of E. lacerata (106 spore/mL) (Table 4). However, wilt biocontrol was not significantly different with higher inoculum concentrations (107 and 108 spore/mL) of the pathogen.
The rhizosphere of healthy plants has been established to harbor richer and more diverse microbial communities than soil associated with the root of diseased plants [27]. Consequently, biocontrol application (such as the addition of B. velezensis and B. thuringiensis) could further increase the microbial diversity in pepper rhizosphere with beneficial strains and shift rhizosphere microbial structure for a more sustainable crop production. The assembly (structure) of microbial community is determined by differential root- exudate production [28]. Rhizosphere microorganisms exist in a complex community, regulated by limiting factors such as space, nutrients and interference-competition mediated by the production of inhibitory metabolites [29]. However, exudates that selectively improve proliferation of pathogen within the rhizosphere will expose host plants to microbial attack [28]. Therefore, introduction of pathogen-suppressive bacterial species (such as antifungal B. velezensis and B. thuringiensis) could check the competitive edge accorded the pathogen by root exudation. In addition to production of bioactive antifungal metabolites, such Bacillus strains could increase microbial diversity in the rhizosphere and promote competition for space and resources (in favour of beneficial microbes).
Contrary to previous reports on improved antimicrobial efficacy associated with the combination of biocontrol agents [30], biocontrol efficacy of antimycotic Bacillus strains EBs02 and EBs04 was not additive. Combining both antifungal strains did not significantly improve wilt severity in infected pepper plants, especially, compared with the biocontrol efficacy of each strain. In a similar report, a collection of Pseudomonas strains was investigated by Vrieze M, et al. [31] for their ability to control Phytophthora infestans, the causal agent of late blight in many economically significant crops. Pseudomonas fluorescens S35 was observed to confer significant protection on three cultivars of potato when applied alone, while its biocontrol potential declined when combined with other strains of Pseudomonas species. Vrieze M, et al. [31] attributed the poor performance of P. fluorescens S35, in dual combination with other strains, to its contrasting mode of action, as well as its inability to compete for space and nutrients.
Conclusion
Emerging infectious diseases of economically significant crops like pepper could cause significant loss to farmers. Wilt caused by E. lacerata has been investigated in tomatoes, while the organism’s pathogenicity in pepper is reported in this study. Previously reported host spectrum of E. lacerata and occurrence of physiological specializations with both medical and agricultural significance make the potential outbreak of Emmia infection threatening. Effect of inoculum dose on the incubation period, disease severity index and biocontrol of wilt caused E. lacerata (in pepper) were investigated.
| Bacillus strain | Inoculum concentration of Emmia species/Wilt severity | ||
|---|---|---|---|
| 106 cfu/mL | 107 cfu/mL | 108 cfu/mL | |
| Bacillus velezensis | 0.75±0.48bc | 1.50±0.65b | 1.13±0.31bc |
| Bacillus thuringiensis | 1.25±0.48b | 1.05±0.65b | 2.25±0.75b |
| B. velezensis + B. thuringiensis | 1.02±0.01bc | 2.00±0.41ab | 1.75±0.48b |
| Negative control | 2.75±0.18a | 2.75±0.42a | 3.25±0.25a |
| Positive control | 0.00c | 0.00c | 0.00c |
Table 3: ** Effect of Bacillus treatment on the severity of pepper wilt.
Negative Control: Infected, untreated plants; Positive control: Uninfected, healthy plants Mean values (± standard error). Means followed by the same letter(s) within a column are not significantly different at 5% probability level according to Duncan’s Multiple Range Test. Table 3: Effect of Bacillus treatment on the severity of pepper wilt.
| Biocontrol treatment/Wilt control (%) | |||
|---|---|---|---|
| Concentration of Emmia species | Bacillus velezensis | Bacillus thuringiensis | B. velezensis + B. thuringiensis |
| 106 spore/mL | 72.75±17.45a | 54.50±17.49a | 64.00±1.02a |
| 107 spore/mL | 47.75±21.80ab | 47.75±21.80a | 29.50±13.14abc |
| 108 spore/mL | 65.25±9.84a | 36.43±18.87a | 45.93±14.69ab |
Table 4: ** Control of pepper wilt with Bacillus strains.
Mean values (± standard error). Means followed by the same letter(s) within a column are not significantly different at 5% probability level according to Duncan’s Multiple Range Test. Table 4: Control of pepper wilt with Bacillus strains.
Increasing the concentration of wilt pathogen to 108 spore/mL did not influence the duration of incubation and wilt severity in infected plants. Bacillus velezensis EBs02 and B. thuringiensis EBs04 inhibited the mycelial growth of E. lacerata and significantly improved wilt severity in infected pepper. However, combining both antifungal Bacillus strains did not potentiate their biocontrol efficacy against Emmia wilt. The susceptibility of solanaceous crops like pepper, consumed as raw or partially cooked diet, to phytopathogenic forms of E. lacerata might be an evolving health concern. Consequently, awareness on the pathogenic and non-pathogenic forms of E. lacerata, as well as early development of sustainable biocontrol measure, such as the application of B. velezensis or B. thuringiensis, is imperative. Also, these strains have exhibited prospects towards management of Emmia wilt and should be investigated for the biocontrol of other wilt-associated plant pathogens, such as Ralstonia solanacearum and Fusarium species. This would significantly improve the yield and quality of susceptible varieties, and equally prompt pepper cultivation, especially in areas endemic to these phytopathogens.
Acknowledgments
Authors acknowledge the support of Bioscience Laboratory of the International Institute of Tropical Agriculture (IITA), Ibadan; as well as the contributions of Organic Farm Unit of the Federal College of Agriculture, Ibadan.
References
-
Adedire OM, Pitan A, Farinu AO, Ogundipe WF (2019) the biocontrol of soil transmitted Cercospora capsici with LactoBacillus plantarum. Journal of Advances in Microbiology 18(3): 1-8.
-
Wang L, Zhong Y, Liu J, Ma R, Miao Y, et al. (2023) Pigment biosynthesis and molecular genetics of fruit color in Pepper. Plants 12(11): 2156.
-
Odo VC, Abu NE, Igu TC, Asogwa LN, Eze CO, et al. (2023) Expression of bioactive compounds in different pepper cultivars (Capsicum annuum L.) in response to different fertilizer treatments. African Journal of Biotechnology 22(8): 153-159.
-
Chen W, Modi D, Picot A (2023) Soil and phytomicrobiome for plant disease suppression and management under climate change: a review. Plants 12(14): 2736.
-
Balamurugan A, Kumar A (2023) Postharvest fruit rot of Bell pepper (Capsicum annuum L.): pathogenicity and host range of Alternaria alternata. Scientia Horticulturae 319: 112156.
-
Xhemali B, Cortiello M, Gjinovci G, Bresilla, B, Stefani E et al. (2023) First report of Colletotrichum scovillei causing anthracnose of pepper in Kosovo. New Disease Reports 47(2): e12189.
-
Pitan AA, Adedire OM, Oluade EA, Adediji DB, Afolabi K, et al. (2020) Incidence, diversity and management of mycopathogens infecting tomato plants in selected Local Government Areas of Oyo State [Unpublished manuscript]. Federal College of Agriculture, Ibadan.
-
Adedire OM, Adedire DE, Ogundipe WF, Farinu AO (2022) Phytopathogenicity and control of Emmia lacerata FMIB29 on tomato (Solanum lycopersicum L.) plant. Archives of Phytopathology and Plant Protection 55(8): 1031-1044.
-
Lee JM, Han E, Kim J, Park JH, Sung GH, et al. (2020) Five Korean cases of respiratory tract infection by filamentous Basidiomycetes. Annals of laboratory medicine 40(1): 84-87.
-
Pan X, Liang J, Zhang J, Liu Y, Liang J, et al. (2022) Emmia lacerata SR5 promotes Michelia macclurei growth by enhancing nutrient uptake and regulating morphology. Journal of Soil Science and Plant Nutrition 22: 985-999.
-
Fusco V, Pasciuta V, Lumia V, Matere A, Battaglia V, et al. (2022) Root and stem rot, and wilting of olive tree caused by Dematophora necatrix and associated with Emmia lacerata in Central Italy. European Journal of Plant Pathology 163: 71-96.
-
Brooks DR, Hoberg EP, Boeger WA, Trivellone V (2022) Emerging infectious disease: An underappreciated area of strategic concern for food security. Transboundary and Emerging Diseases 69: 254-267.
-
Adedire DE, Owoeye TE, Farinu OM, Ogundipe WF, Adedire OM (2023) Management of fusarium wilt disease of maize with selected antimycotic plant extracts and inhibitory Bacillus strains. Current Microbiology 80: 204.
-
Chatterton S, Schwinghamer TD, Page A, Davidson RB, Harding MW (2023) Inoculum dose–disease response relationships for the pea root rot pathogen, Aphanomyces euteiches, are dependent on soil type and other pathogens. Frontiers in Plant Science 14: 1115420.
-
Bindal S, Sheu ZM, Kenyon L, Taher D, Rakha M (2023) Novel sources of resistance to fusarium wilt in Luffa species. Frontiers in Plant Science 14: 1116006.
-
Cao Y, Hualiang P, Chandrangsu P, Yongtao L, Yuqi W, et al. (2018) Antagonism of two plant-growth promoting Bacillus velezensis isolates against Ralstonia solanacearum and Fusarium oxysporum. Scientific Reports 8: 4360-4368.
-
Hammami R, Oueslati M, Smiri M, Nefzi S, Ruissi M, et al. (2022) Epiphytic yeasts and bacteria as candidate biocontrol agents of green and blue molds of Citrus fruits. Journal of Fungi 8(8): 818.
-
Jaiswal AK, Frenkel O, Elad Y, Lew B, Graber ER (2015) Non-monotonic influence of biochar dose on bean seedling growth and susceptibility to Rhizoctonia solani: the Shifted Rmax-Effect. Plant Soil 395: 125-140.
-
Yin J, Bai R, Yuan L, Huang J (2023) Application of Ceriporia lacerata HG2011 as biocontrol agent against multiple phytopathogenic fungi and oomycetes. Pesticide Biochemistry and Physiology 190: 105316.
-
Mehrabi R, Bahkali AH, Elsalam KA, Moslem M, barek BS, et al. (2011) Horizontal gene and chromosome transfer in plant pathogenic fungi affecting host range. FEMS Microbiology Review 35(3): 542-54.
-
Friesen TL, Stukenbrock EH, Liu ZH, Meinhardt S, Ling H, et al. (2006) Emergence of a new disease as a result of interspecific virulence gene transfer. Nature Genetics 38: 953-956.
-
Ma LJ, Does HC, Borkovich KA, Jeffrey JC, Marie JD, et al. (2010) Comparative genomics reveals mobile pathogenicity chromosomes in Fusarium. Nature 464: 367-373.
-
Enagbonma BJ, Fadiji AE, Ayangbenro AS, Babalola OO (2023) Communication between plants and rhizosphere microbiome: exploring the root microbiome for sustainable agriculture. Microorganisms 11(8): 2003.
-
Jan F, Arshad H, Ahad M, Jamal A, Smith DL (2023) In vitro assessment of Bacillus subtilis FJ3 affirms its biocontrol and plant growth promoting potential. Frontiers in Plant Science 14: 1205894.
-
Sicuia OA, Toma RC, Diguta CF, Matei F, Cornea CP (2023) In vitro evaluation of some endophytic Bacillus to potentially inhibit Grape and Grapevine fungal pathogens. Plants 12(13): 2553.
-
Fatima R, Mahmood T, Moosa A, Aslam MN, Shakeel MT, et al. (2023) Bacillus thuringiensis CHGP12 uses a multifaceted approach for the suppression of Fusarium oxysporum f. sp. ciceris and to enhance the biomass of chickpea plants. Pest Management Science 79(1): 336- 348.
-
Zhu Z, Wu G, Deng R, Xiaoying HU, Yaping C (2023) spatiotemporal biocontrol and rhizosphere microbiome analysis of Fusarium wilt of banana. Communications Biology 6: 27.
-
Li M, Song Z, Li Z, Qiao R, Zhang P, et al. (2022) Populus root exudates are associated with rhizosphere microbial communities and symbiotic patterns. Frontiers in Microbiology 13: 1042944.
-
Chepsergon J, Moleleki LN (2023) Rhizosphere bacterial interactions and impact on plant health. Current Opinion in Microbiology 73: 102297.
-
Bonaterra A, Badosa E, Daranas N, Frances J, Rosello G, et al. (2022) Bacteria as biological control agents of plant diseases. Microorganisms 10(9): 1759.
-
Vrieze M, Germanier F, Vuille N, Weisskopf L (2018) Combining different potato-associated Pseudomonas strains for improved biocontrol of Phytophthora infestans. Frontiers in Microbiology 9: 2573.
- Acido Labile or Gastro Irritant Apis and Enteric Release in Galenic Practice: An Overview
- A Study on Knowledge, Attitude and Practice of Hand Hygiene among Healthcare Professionals at a Tertiary Care Hospital, India
- Vanilla’s Chemistry
- Marine Anti-Cancer Compounds and Adverse Effects of Global Warming on Oceans: An Overview
- Serological Investigation of Chikungunya Virus Antibody among Malaria-Suspected Febrile Patients in Some Healthcare Facilities in Rivers State
- Pharmaceutical Applications of White Button Mushroom (Agaricus Bisporus)