Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
Our study using the comet assay to detect genotoxicity of carcinogens in multiple mouse organs reveled that the correlation between genotoxicity and carcinogenicity on an organ-by-organ basis is not always high. One reason for this discrepancy is thought to be the difference in the administration method; carcinogenicity is detected by long-term administration at low doses, while genotoxicity has been detected by a single dose in the high dose range known as the maximum tolerated dose (MTD) using the comet assay. If there is such a large discrepancy between the results of in vivo genotoxicity and long-term carcinogenicity tests, it would be difficult to judge whether the experimentally suggested carcinogenicity is due to a genotoxic mechanism. Here, we compared the results of comet assay when carcinogens targeting and not targeting rodent liver were given by feeding or drinking with those when gavaged and examined the consistency with the results of carcinogenicity tests. Although single gavage of carcinogens targeting rodent liver at daily dose used in the carcinogenicity study induced DNA damage not only in the liver (carcinogenicity target organ) but also organs other than the liver, their triple gavages induced DNA damage only in the liver. Furthermore, their feeding and drinking for 4- and 6-days induced DNA damage in the carcinogenesis target organs. Their feeding and drinking for 1- and 2-days sometimes induced DNA damage in carcinogenesis not-targeting organs. Good correlation was shown between carcinogenicity targeting the liver and increase of their genotoxicity in the liver dependent to the duration of feeding and drinking period when they were given to mice continuously in the diet or drinking water. Therefore, the comet assay by continuous dosing of chemicals in the diet and drinking water would give useful information to predict carcinogenicity targeting sites.
Yokohama N¹, Kawaguchi S², Yamamoto A², Nakamura T³, and Sasaki Yu F²,³
¹Advanced Material and Biological Engineering Course, National Institute of Technology, Hachinohe College, Japan ²Material and Biological Engineering Course, National Institute of Technology, Hachinohe College, Japan ³Department of Pharmaceutical Health Care, Faculty of Pharmaceutical Sciences, Himeji Dokkyo University, Himeji, Japan
Abbreviations
MTD: Maximum Tolerated Dose; DMN: Dimethylnitrosamine; DEN: Diethylnitrosamine; DAB: p-Dimethylamonoazobenzene; LGT: Low Melting Point; MMS: Methanesulfonate; EMS: Ethyl Methanesulfonate; EAFUS: Everything Added to Food in the United States.
Introduction
We have evaluated the genotoxicity of 208 compounds selected from chemicals selected from the IARC Monographs and U.S. NTP carcinogenicity database in multiple mouse organs using the comet assay and have clarified the correlation between genotoxicity and carcinogenicity on an organ-by- organ basis [1, 2]. As a result, it has already been reported that when a compound is positive in the comet assay in at least one organ, the positive and negative rates are in good agreement with the presence or absence of its carcinogenicity. However, the organs in which genotoxicity is observed are not always the target organs for carcinogenicity, and the correlation between genotoxicity and carcinogenicity on an organ-by- organ basis is not always high, and the correlation differs greatly depending on the organ type. One reason for this is thought to be the difference in the administration method; carcinogenicity is detected by long-term administration at low doses, while genotoxicity has been detected by a single dose in the high dose range known as the maximum tolerated dose (MTD) using the comet assay. Traditionally, in vivo genotoxicity tests have often been performed by single intraperitoneal or gavage administration at MTD, and our in vivo comet assay data were obtained in this way. However, carcinogenicity tests often detect carcinogenicity by long- term administration in the diet. It has also been reported that in carcinogenicity tests, the positive rate is lower when animals are fed in diet or drinking water than when they receive gavage administration [3]. This suggests that the form of administration also has a large effect on genotoxicity. If there is such a large discrepancy between the results of in vivo genotoxicity and long-term carcinogenicity tests, it would be difficult to judge whether the experimentally suggested carcinogenicity is due to a genotoxic mechanism. Nevertheless, to date, genotoxicity has rarely been studied through feeding or drinking. The purpose of this study was to explore one of the causes of the large discrepancy between the results of in vivo genotoxicity by comet assay and long- term carcinogenicity tests for rodent hepatic carcinogens. Here, we compared the results of comet assay when carcinogens targeting and not targeting rodent liver were given by feeding or drinking with those when gavaged and examined the consistency with the results of carcinogenicity tests.
Materials and Methods
Chemicals and Animals
Tested carcinogens were divided into two categories, i.e., carcinogens targeting rodent liver and those targeting rodent organs other than the liver. The former includes dimethylnitrosamine (DMN), diethylnitrosamine (DEN), p-dimethylamonoazobenzene (DAB), 2,4-diaminotoluene (2,4DAT), IQ, and 1,2-dimethylhydrazine (1,2DMH) and the latter includes azobenzene, aniline hydrochloride, and PhIP. Their carcinogenesis targeting organs and dose used in the carcinogenicity test were shown in Table 1.
Regular (GP-42) and low melting point (LGT) agarose were obtained from Nacalai Tesque (Kyoto) and diluted, respectively, to 1% and 2% in physiological saline. Male ddY mice were obtained from Japan SLC Co at 7 weeks of age and used after 1 week of acclimatization. They were fed commercial powdered basal diet (CRF-1, Oriental Yeast Industries Co., Tokyo, Japan) and tap water ad libitum throughout the acclimatization period and the experiment. The animal room was at 20 - 24 ℃ with a 12-h light-dark cycle.
The Comet Assay
The experiment followed the design of our previous comet assay studies on multiple organs [4, 5, 6]. Groups of 4 male ddY mice were given drinking water containing DMN, DEN, 1,2DMH, and aniline hydrochloride or fed a diet containing azobenzene, DAB, 2,4DAT, IQ, PhIP, for up to 6 days at the dose levels used in carcinogenicity study (mean body weight at starting was3 7.1g). Slides for comet observation were prepared after feeding or drinking for 1, 2, 4, and 6 days. Feeding or drinking started at 9:00a.m. and daily diet intake, drinking water intake, and body weight were measured at every 9:00a.m. During the feeding and drinking period, mice were carefully observed for pharmacotoxic signs. Mice were sacrificed at around 10:00 a.m. by bleeding under anesthesia after feeding or drinking for 1, 2, 4, and 6 days and eight organs (glandular stomach, colon, liver, kidneys, urinary bladder, lungs, brain, and bone marrow) were removed from each animal, namely, and used for the comet assay [3]. When necropsies were performed, each organ was examined for changes in size, color, and texture and a small portion of each organ was fixed in 10% formaldehyde, dehydrated, and embedded in paraffin and subjected to histopathological examination.
In another experiment, carcinogens dissolved in vehicles shown in Table 1 were administered orally to group of 4 mice per group were treated with each carcinogen by giving a single gavage at daily dose in carcinogenicity study and triple gavages with the interval of 24 h at daily dose in carcinogenicity study. Also, they were treated with each carcinogen by giving a single gavage at MTD level. The administration volume was 10 mL/kg. From shortly after they were treated until just before they were sacrificed, mice were carefully observed for pharmacotoxic signs. Mice were sacrificed 3 and 24 hours after the final gavage, and then eight organs, (glandular stomach, colon, liver, kidney, bladder, lung, brain, and bone marrow) were removed from each individual and subjected to comet assay [1, 2].
The liver, kidney, lung, and brain were minced, suspended in 4 mL chilled homogenizing solution (pH 7.5) containing 0.075 M NaCl and 0.024 M Na2EDTA, and then homogenized gently using a Potter-Elvehjem type homogenizer at 500 - 800 rpm, in ice [1, 4, 5, 6].
| Chemicals | Vehiclea | Dose for feeding or drinking (ppm) b | Mean daily intake | Mean weight gain | Carcinogenesis target organc | LD50d | IARCe | Ref f | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Chemical (mg/ kgBW/ day) | Water (mL/ mouse/ day) | Diet (g/ mouse/ day) | (g/ mouse) | |||||||
| Control | - | 0 | 0 | 3.9 | 4.8 | 3.45 | - | - | - | - |
| Azobenzene | O | 400 | 44.3 | - | 3.7 | 4.3 | - | 500 | 3 | 16 |
| Aniline hydrochloride | S | 12000 | 1340 | 3.9 | - | 3.83 | - | 464 | 3 | 17 |
| IQ | O | 300 | 32.6 | - | 3.8 | 3.5 | L | 400 | 2A | 18 |
| PhIP | O | 400 | 48 | - | 4.2 | 3.22 | Sp | <150 | 2B | 19 |
| 2,4DAT | O | 2B200 | 22.3 | - | 3.9 | 3.56 | L | 350 | 2B | 20 |
| DAB | O | 560** | 56 | - | 3.5 | 3.64 | L** | 200* | 2B | 10 |
| DMN | S | 10 | 1.11 | 3.9 | - | 3.26 | L | 37* | 2A | 21 |
| DEN | S | 420 | 52.8 | 4.2 | - | 4.12 | L | 200 | 2A | 22 |
| 1,2DMH | S | 10 | 1.11 | 3.9 | - | 2.98 | L,K,Co | 26 | 2A | 9 |
Table 1: Daily intake of studied carcinogens, drinking water, diet intake, and weight gain in mice dosed by feeding and drinking.
*Because data of acute toxicity data in mice by single oral gavage are not available, data of acute toxicity data in rats by single oral gavage are shown. **Because carcinogenesis study data by feeding in mice are not available, data of carcinogenesis study data in rats by feeding are shown. a Vehicle for gavage administration; O, Olive oil; S, saline b Dose used in carcinogenesis study in mice by feeding or drinking c Carcinogenesis target organs in mice by feeding or drinking; L, liver; K, kidney; Sp, spleen; d LD50 by single oral gavage in mice e IARC classification, probable; 2A, Probably carcinogenic to humans; 2B, Possibly carcinogenic to humans; 3, Not classifiable as to its carcinogenicity to humans fReferences for carcinogenesis target organs in mice and LD50 in mice.
The glandular stomach, colon, and urinary bladder were opened and rinsed with physiological saline; then the mucosa was scraped into 4 ml chilled homogenizing buffer and homogenized gently using a Potter-Elvehjem type homogenizer at 500 - 800 rpm, in ice. To obtain nuclei, the homogenate was centrifuged at 700g for 10 min at ℃, and the precipitate was re-suspended in chilled homogenizing buffer at 1 g organ weight per milliliter. Seventy-five µL agarose GP- 42 was quickly layered on a slide (Matsunami Glass Ind., Ltd., Osaka, Japan) coated with agarose GP-42 and covered with another slide. The slide sandwiches were placed horizontally to allow the agarose to solidify. The nucleus suspension was mixed 1:1 (v/v) with 2%, 45℃, agarose-LGT, and 75 µL of the nucleus mixture was quickly layered in the same manner after removal of the covering slide. Finally, 75 µL of agarose GP-42 was quickly layered on again. Slides prepared from nuclei isolated by homogenization were placed in a chilled lysing solution (2.5 M NaCl, 100 mM Na2EDTA, 10 mM Trizma, 1% sarkosyl, 10% DMSO, and 1% Triton X-100, pH 10) [4, 5, 6] and kept at 0OC in the dark for about one night, then in chilled alkaline solution (300 mM NaOH and 1 mM Na2EDTA, pH 13) for 10 min in the dark at 0oC [4, 5, 6]. Electrophoresis was conducted at ℃ in the dark for 15 min at 25 V (0.96 V/ cm) and approximately 250 mA. The slides were neutralized and then stained with 50 µL of 20 µg/ml ethidium bromide (Wako Pure Chemical Industries, Ltd.) [4, 5, 6]. We examined and photographed 50 nuclei per slide at 200 x magnification with the aid of a fluorescence microscope. The length of the whole comet (“length”) and the diameter of the head (“diameter”) were measured for 50 nuclei per organ per animal. We calculated tail length as the difference between length and diameter for each of 50 nuclei. Mean migration of 50 nuclei from each organ was calculated for each individual animal. The differences between the averages of four treated animals and the untreated control animals were compared with the t test. A p-value less than 0.05 was considered statistically significant.
Results
The results by gavage administration and feeding (drinking) were shown in Figures 1 & 2, respectively. No death, morbidity, or clinical signs were observed after any single gavage administration. Remarkable decreases in diet, water intake, and body weight gain were not observed in the feeding period. Necropsy and histopathological examination of tissue sections stained by the hematoxylin-eosin revealed no treatment effect on any organ examined. Thus, any DNA damage observed was not likely to be due to general cytotoxicity (necrosis) and apoptosis.
Carcinogens Targeting Rodent Liver
Single gavage of DMN at daily dose used in the carcinogenicity study (1 mg/kg) and MTD level (50 mg/kg) increased tail length in the stomach, colon, kidney, urinary bladder, lung, and bone marrow, but its triple gavage did not increase tail length at any organs studied (Figure 1A). Drinking of DMN increased tail length in the stomach only after its 4-day drinking period and liver after its 1-, 2-, 4-, and 6-day drinking period (Figure 2A). Tail length in the liver increased dependently to the duration of drinking period (Pearson’s correlation coefficient: 0.9496).
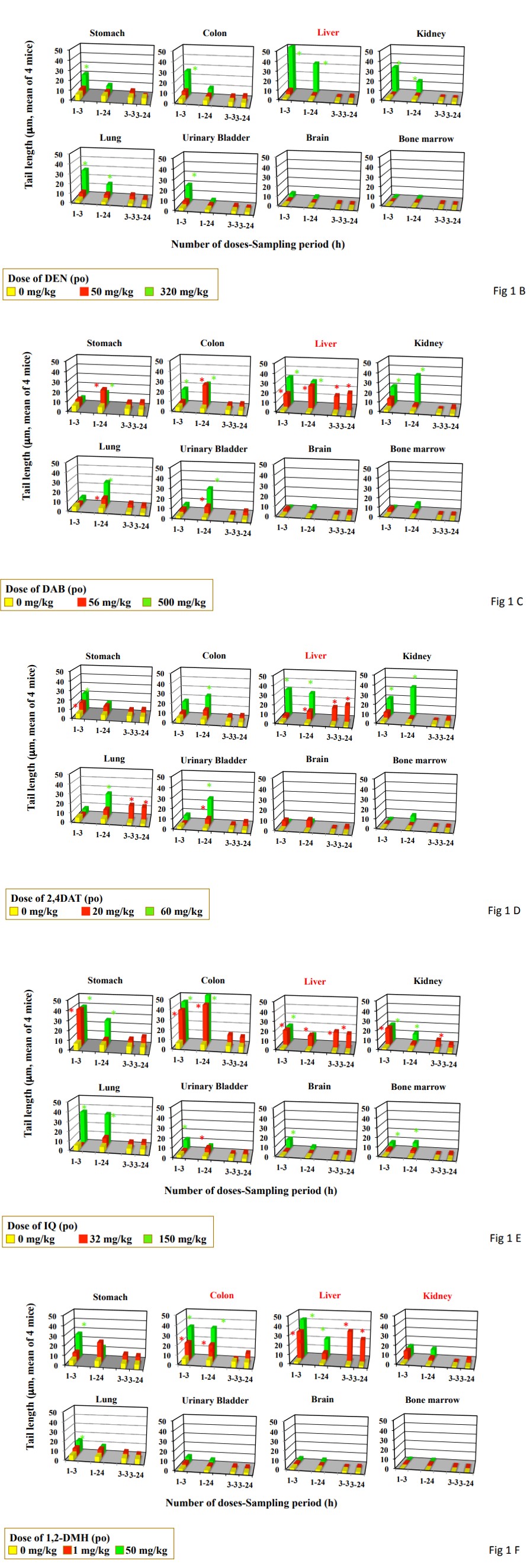
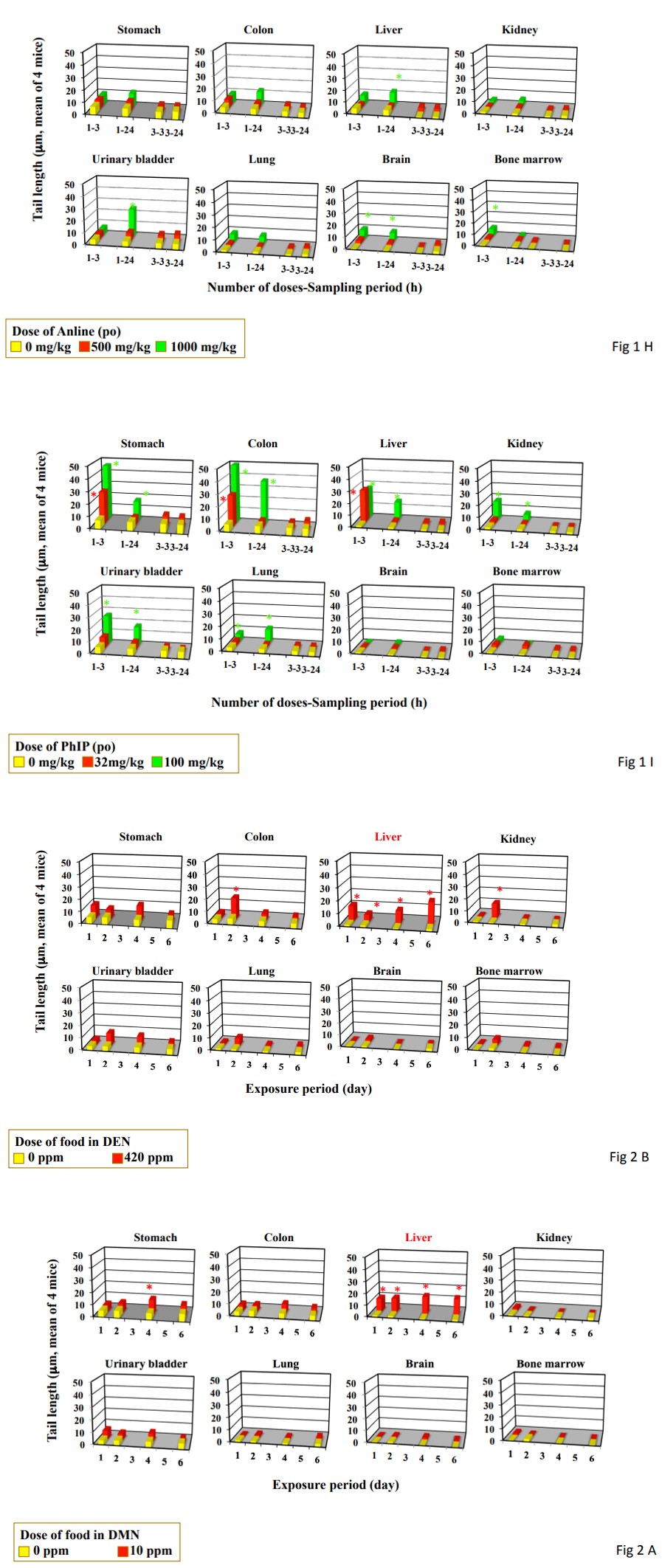
Figure 1: DNA damage measured by the comet assay in the organs from mice treated by single and triple gavages. Data represent mean of 4 mice. Statistical difference from untreated control: *p<0.05. Organs in red are carcinogenesis targeting organ in mice by feeding or drinking. A, DMN; B, DEN; C, DAB; D, 2,4DAT; E, IQ, F, 1,2DMH; G, azobenzene; H, aniline hydrochloride; I, PhIP.
Single gavage of DEN at MTD level (320 mg/kg) increased tail length in the studied organs except for brain and bone marrow, but its single gavage at daily dose used in the carcinogenicity study (50 mg/kg) did not increase tail length in any studied organs (Figure 1B). Its triple gavages also did not increase tail length in any studied organs. Drinking of
DEN increased tail length in the liver after its 1-, 2-, 4-, and 6-day drinking at the dose used in the carcinogenicity study (420 ppm). Tail length in the colon and kidney increased only after 2-day drinking (Figure 2B). Tail length in the liver increased dependently to the duration of drinking period (Pearson’s correlation coefficient: 0.9829).
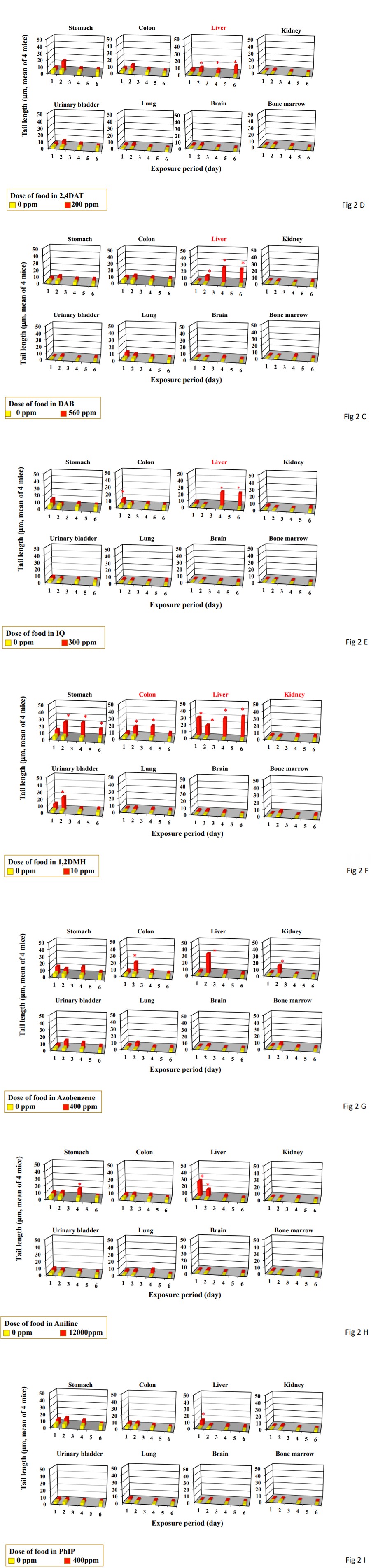
Figure 2: DNA damage measured by the comet assay in the organs from mice treated by feeding and drinking for up to 6 days. Data represent mean of 4 mice. Statistical difference from untreated control: *p<0.05. Organs in red are carcinogenesis targeting organ mice by feeding or drinking. A, DMN; B, DEN; C, DAB; D, 2,4DAT; E, IQ, F, 1,2DMH; G, azobenzene; H, aniline hydrochloride; I, PhIP.
Single gavage of DAB at daily dose used in the carcinogenicity study (56 mg/kg) increased tail length in the studied organs except for the kidney, brain and bone marrow (Figure 1C). Its triple gavages at 56 mg/kg increased tail length only in the liver. Feeding of DAB at the dose used in the carcinogenicity study (560 ppm) increased tail length in the liver after its 2-, 4-, and 6-day feeding (Figure 2C). The tail length in the liver increased dependently to the duration of feeding period (Pearson’s correlation coefficient: 0.809).
Single gavage of 2,4DAT at daily dose used in the carcinogenicity study (20 mg/kg) increased tail length in the stomach, liver, urinary bladder, and lung and at MTD level (60 mg/kg) in the stomach, colon, liver, kidney, urinary bladder, and lung (Figure 1D). Its triple gavages at 20 mg/kg increased tail length in the liver and urinary bladder. Feeding of 2,4DAT at the dose used in the carcinogenicity study (200 ppm) increased tail length in the liver after its 2-, 4-, and 6-day feeding (Figure 2D). Tail length increased dependently to and the duration of feeding period (Pearson’s correlation coefficient: 0.853).
Single gavage of IQ at daily dose used in the carcinogenicity study (30 mg/kg) increased tail length in the stomach, colon, and liver, kidney, and urinary bladder, and at NTD level (150 mg/kg) in all the organs studied (Figure 1E). Its triple gavages at 32 mg/kg increased tail length in the liver and kidney. Feeding of IQ at the dose used in the carcinogenicity study (300 ppm) increased only in the liver after its 4- and 6-day feeding (Figure 2E). Tail length in the liver increased dependent to the duration of feeding period (Pearson’s correlation coefficient: 0.8500) Although single gavage of 1,2DMH at daily dose used in the carcinogenicity study (1 mg/kg) increased tail length in the colon, and liver, and kidney, its triple gavages at 1 mg/kg increased tail length only in the liver (Figure 1F). Drinking of 1,2DMH at the dose used in the carcinogenicity study (10 ppm) increased tail length in the stomach, liver, and urinary bladder after its 4-day drinking. After 6-day drinking, tail length increased in the stomach and liver (Figure 2F). After 2-day drinking, tail length increased in the stomach, colon liver, and urinary bladder. After 1-day drinking, tail length increased only in the liver. Tail length in the liver increased dependently to the duration of drinking (Pearson’s correlation coefficient: 0.9573).
Carcinogens not Targeting Rodent Liver
Single and triple gavages of azobenzene at daily dose used in the carcinogenicity study (44 mg/kg) did not increase tail length at any studied organs (Figure 1G). Its single gavage at MTD level (500 mg/kg) increased tail length in the stomach, colon, and urinary bladder. Its drinking at the dose used in the carcinogenicity study (400 ppm) increased tail length in the colon, liver, and kidney only after its 2-day feeding (Figure 2G). The tail length in the liver decreased dependent to the duration of drinking (Piason’s correlation coefficient: -0.885) Single and triple gavages of aniline hydrochloride at daily dose used in the carcinogenicity study (1000 mg/kg) did not increase tail length at any studied organs except for urinary bladder (Figure 1H). Only in the urinary bladder, tail length increased by its single gavage. Its drinking at the dose used in the carcinogenicity study (12000 ppm) increased tail length in the stomach only after 4 day-drinking and in the liver after its 1- and 2-day feeding (Figure 2H). The tail length in the liver decreased dependently to the duration of drinking (Piason’s correlation coefficient: -0.9251) Single gavages of PhIP at daily dose used in the carcinogenicity study (32 mg/ kg) increased tail length in the stomach and colon, but in the stomach, colon, liver, kidney, urinary bladder, and lung at MTD level (100 mg/kg) (Figure 1I). On the other hand, its triple gavages at 32 mg/kg did not increase tail length in any studied organs. Its feeding at the dose used in the carcinogenicity study (400 ppm) did not increase tail length in any studied organs (Figure 2I).
Discussion
Although single gavage of carcinogens targeting rodent liver at daily dose used in the carcinogenicity study induced DNA damage not only in the liver (carcinogenicity target organ) but also organs other than the liver, their triple gavages induced DNA damage only in the liver. Furthermore, their feeding and drinking for 4- and 6-days induced DNA damage in the liver and other carcinogenesis target organs, i.e., colon and kidney for 1,2DMH, except for the stomach for 1,2DMH. Their feeding and drinking for 1- and 2-days sometimes induce DNA damage in carcinogenesis not-targeting organs. When carcinogens targeting to the liver were given to mice continuously in the diet or drinking water, tail length in the liver increased dependently the duration of feeding period. On the contrary, carcinogens not targeting the liver did not increase the tail length in the liver dependently to the duration of feeding period.
From the above, the possibility would be considered that the induction of DNA damage in carcinogenicity not-targeting organs for by feeding and drinking administration is limited to the early stages of feeding and drinking, and conversely, organs in which the induction of DNA damage persists until 4- and 6-days may become target organs for carcinogenicity.
Although single gavage of DAB, an azo dye targeting the liver, induced DNA damage not only in the live but also various carcinogenesis not targeting organs (stomach, colon, kidney, urinary bladder, and lung), its triples gavages induced DNA only in the liver. One possible explanation for this disappearance in the organs other than the liver might be that their triple gavages lower the activity of azo reduction in the GI-tract and/or the sensitivity of mucosa cells of GI-tract against azo metabolites. In the contrast to their genotoxicity in GI-tract, DAB yielded DNA damage in the liver after single and triple gavages, which might suggest that azo reduction occurred after triple gavages. Therefore, the former possibility would not be plausible.
The latter possibility might be explained by the lowering of sensitivity in non-targeting organs against carcinogens due to an elevation of the removal of DNA damages after multiple dosing. The elevation of the DNA repair activity, adaptive, response, is well known in bacteria and in vitro eukaryotes after conditioning to low dose of ionizing radiation and some alkylating agents. In mice treated with 150 mg/kg methyl methanesulfonate (MMS), the yield of chromosomal aberrations in bone marrow cells reduced when they were pre-treated with MMS at 50 mg/kg [7]. The clastogenic effect of ethyl methanesulfonate (EMS) at 240 mg/kg was also reduced by low conditioning dosing of EMS at 80 mg/ kg prior to its challenging dosing [8]. It is discussed that the low dose of alkylating agents employed has made the cells less sensitive against further clastogenic effect of challenge dose of alkylating agents and the phenomenon of adaptive response to alkylating agents can be encountered in in vivo mammalian cells [7, 8]. Considering those results, it might be possible that the removal of DNA damage produced by azo metabolites is elevated by their multiple dosing in cells in the organs other than the liver. To determine whether the latter possibility is valid, however, mechanistic studies, such as the differences in adaptive repair between carcinogenesis target and non-target organs, would be necessary. 1,2DMH is carcinogenic to mouse colon and liver [9] and DAB is carcinogenic to mouse liver [10]. After triple dosing, DAB yielded DNA damage in the liver and DMH in the colon and liver. Therefore, the correlation of their carcinogenicity and genotoxicity is better when they were given by triple than single gavage.
When 1,2DMH, and DAB were given mice by feeding or drinking, their genotoxicity in the liver increased dependently to the duration of feeding or drinking period. This increase with an administration period parallels with the observation that their genotoxicity was observed after not only single but also triple gavages. On the contrary, when azobenzene and aniline, that are carcinogens not targeting the liver, were given mice by feeding or drinking, their genotoxicity decreased dependently to the duration of feeding or drinking period. Animals fed diet containing chemicals consume less food than they would if the chemicals were absent thus take in a smaller amount of chemical than planned [11]. Decreases in food (caloric) intake that are often associated with reductions in tumor burden and improvements in survival might affect genotoxicity [11]. In this study, any decreases in food and water intake and body weight gain were not observed in the feeding period. Gavage-administered carcinogens tended to be more genotoxic, which may be due to that gavage- administered carcinogens may tend to saturate metabolic/ detoxification pathway like as Yuan, et al. [12] showed in their study with benzyl acetate.
Although 1,2DMH given by gavage once per week induces tumors in mouse colon, anal region, liver, and uterus, the incidence of tumors in the colon and anal region was not observed when it was given continuously in the drinking- water [9]. In the mucosa of GI-tract cells having DNA damage induced in early stages of drinking might diminish due to proliferation of mucosa cell, which might prevent the accumulation of cells having DNA damage until late stage of drinking. Therefore, it could be explained the reason why DNA damage in the colon that is one carcinogenesis target organ did not increase dependently to the duration of drinking period. Therefore, our present result suggested good correlation between carcinogenicity of 1,2DMH given continuously in the drinking-water and increasing tendency of genotoxicity with dinking period in its target organ (liver) except for the GI-tract (colon). At the International Workshop on Genotoxicity Test Procedures (IWGTP) in 1999 [13] an expert panel met to develop guidelines for the use of the comet assay in genetic toxicology. In a consensus of minimal standards for obtaining reproducible and reliable comet data deemed suitable for regulatory submission, it is recommended that test substances are generally administered as a single treatment at MTD level which should be sufficient high to elicit signs of toxicity. The gavage route may bear little, if any, resemblance to human exposures to chemicals used as food additives [14]. Gavage is appropriate for testing orally administered drugs [3]. In the U.S.A., the more than 3000 total substances together comprise an inventory often referred to as “Everything” Added to Food in the United States (EAFUS) [15]. The distribution of EAFUS rodent carcinogenesis and non-carcinogenesis revealed 17 noncarcinogens and 1 carcinogen among chemicals administered in the feed, compared with 7 noncarcinogens and 16 carcinogens among gavage-administered chemicals [3]. Thus, the distribution clearly shows that noncarcinogens are significantly more prevalent in feed studies compared with gavage studies and that the use of non-feed routes may tend to increase the likelihood of a positive rodent cancer- test response [3]. Saturation of metabolic/detoxification pathway may explain why gavage-administered chemicals generally tended to be more carcinogenic [3]. However, for real-life dose levels in humans, saturation may seldom occur [3]. Like as in rodent cancer-test response, present results showed that gavage-administered carcinogens targeting rodent liver tended to be genotoxic in carcinogenesis not- targeting organs as well as the liver. If higher response of their genotoxicity in gavage studies could be explained by saturation of metabolic/detoxification pathway, genotoxic response by continuous dosing might be more meaningful toxicologically. The comet assay by continuous dosing of chemicals in the diet and drinking water would give useful information to predict carcinogenicity targeting sites.
Acknowledgements
This research was conducted in Advanced Material and Biological Engineering Course, National Institute of Technology, Hachinohe College as part of a graduation research project by Natsue Yokohama under the research guidance of course staffs based on allocation of school educational expenses. The authors acknowledge the Advanced Material and Biological Engineering Course, National Institute of Technology, Hachinohe College.
References
-
Sasaki YF, Sekihashi, K, Izumiyama F, Nishidate E, Saga A, et al. (2000) The comet assay with multiple mouse organs: Comparison of comet assay results and carcinogenicity with 208 chemicals selected from the IARC Monographs and U.S. NTP carcinogenicity database. CRC Crt Rev Toxicol 30(6): 629-799.
-
Sasaki, Y.F., S. Kawaguchi, A. Kamaya, M. Ohshita, K. Kabasawa, K. Iwama, K. Taniguchi and S. Tsuda (2002) The comet assay with eight mouse organs: Results with 39 currently used food additives, Mutation Res 519(1-2): 103-119.
-
Johanso FM (2002) How many food additives are rodent carcinogens? Environ Molecular Mutagenesis 39(1): 69- 80.
-
Fairbairn DW, Olive PL, O’Neill KL (1995) The comet assay: a comprehensive review. Mutation Res 339(1): 37-59.
-
Sasaki YF, Tsuda S, Izumiyama F, Nishidate E (1997) Detection of chemically induced DNA lesions in multiple mouse organs (liver, lung, spleen, kidney, and bone marrow) using the alkaline single cell gel electrophoresis (Comet) assay. Mutation Res 388(1): 33-44.
-
Kawaguchi S, Nakamura T, Honda G, Yokohama N, Sasaki YF (2008) In vivo genotoxic potential of kojic acid in rodent multiple organs detected by the comet assay. G&E 30(1): 25-32.
-
Mahmood R, Vasudev V, Harish SK, Guruprasad KP (1996) Inducible protective processes in animal systems: adaptive response to a low dose of methyl methanesulfonate in mouse bone marrow cells. Indian J Exp Biol 34(6): 502-507.
-
Mahmood R, Vasudev V (1993) Inducible protective processes in animal systems: IV. Adaptation of mouse bone marrow cells to a low dose of ethyl methanesulfonate. Mutagenesis 8(1): 83-86.
-
IARC (1999) Re-evaluation of some organic chemicals, hydrazine, and hydrogen peroxide: IARC Momographs on the Evaluation of the Carcinogenic Risk to Humans 71: 947-989.
-
IARC (1975) p-Dimethylaminoazobenzene: IARC Monographs on the Evaluation of Carcinogenesis Risk of Chemicals to Man. France, Lyon, 8: 125-139.
-
Haseman JK, Johanson FM (1996) Analysis of National Toxicology Program rodent bioassay data for anticarcinogenic effects, Mutation Res 350(1): 131-141.
-
Yuan JH, Goel TJ, Abdo K, Clark J, Espinosa O, et al. (1995) Effects of gavage versus dosed feed administration on the toxicokinetics of benzyl acetate in rats and mice. Food Chem Toxicol 33: 151-158.
-
Tice RR, Agurell E, Anderson D, Burlinson B, Hartmann A, et al. (2000) Single cell gel/comet assay: guidelines for in vitro and in vivo genetic toxicology testing. Environ Mol Mutagen 35(3): 206-221.
-
Weil CS (1972) Guideline for experiments to predict the degree of safety of a material for man. Toxicol Appl Pharmacol 21(2): 194-199.
-
(2000) Everything Added to Food in the United States (EAFUS). Food and Drug Administration (FDA).
-
National Cancer Institute (1979) Bioassay of azobenzene for poissible carcinogenicity. CAS No. 103-33-3, Technical Report Series No.154.
-
National Cancer Institute (1978) Bioassay of aniline hydrochloride for possible carcinogenicity. CAS No. 142- 04-1, Technical Report Series No.130.
-
Hoshi M, Wanibuchi H, Salim EI, Morimura K, Murai T, et al. (2004) Carcinogenic Potential of 2-Amino-3- methylimidazo [4,5-f]quinoline (IQ) in Severe Combined Immunodeficient (SCID) Mice. J Toxicol Pathol 17(1): 17- 23.
-
IARC (1975) PhIP (2-amino-1-methyl-6- phenylimidzo[4,5-b] pyridine: IARC Monographs on the Evaluation of Carcinogenesis Risk of Chemicals to Man 56: 229-242.
-
Sontag JM (1981) Carcinogenicity of substituted- benzenediamines (phenylenediamines) in rats and mice. J Natl Cancer Inst 66(3): 591-602.
-
Griciute LM, Castegnaro M, Bereziat JC (1981) Influence of ethyl alcohol on carcinogenesis with N-nitrosodimethylamine. Cancer Lett 13(4): 345-352.
-
Takayama S, Ohta K (1965) Induction of malignant tumors in various strains of mice by oral administration of N-nitrosodimethylamine and N-nitrosodiethylamine. Gann 56(2): 189-199.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells
- The Role of Faculty in Assessment: Traditional vs. Competency- Based Medical Education in Toxicology Learning