Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
Among currently used food dyes, we have reported that four azo dye and three xanthene dyes that were given to mice by single gavage at a low dose (10 or 100 mg/kg) induced DNA damage in mouse gastrointestinal tract (GI-tract). When the susception of carcinogenicity by genotoxic mechanism cannot be excluded, ADI cannot be set for food additives, pesticides, and veterinary drugs. Therefore, it is serious issues whether observed their genotoxicity in mouse GI-tract lead to their carcinogenicity. Although carcinogenicity is examined by long-term administration in feed or drinking water, in vivo genotoxicity tests have been traditionally performed by single intraperitoneal or gavage administration at MTD. Here, we examined the genotoxicity of food dyes given continuously by drinking, to re-evaluate DNA damage induced by food dyes given by gavage in mouse GItract. Although single gavage of food dyes at 2000 mg/kg induced DNA damage in GI-tract, their administration by continuous drinking at daily intake higher than 2000 mg/kg did not induce DNA damage in GI-tract. Based on our previous discussion that the results from continuous administration by drinking or feeding are likely to reflect carcinogenicity by genotoxic mechanisms better than the results from single gavage administration, in spite of the presence of positive responses in mouse GI-tract by single gavage of food dyes, the absence of positive response in mouse GI-tract by continuous drinking administration of food dyes could be considered to show the absence of carcinogenicity by genotoxic mechanisms.
Yokohama N¹, Zenke A², Kawaguchi S², Nakamura T³, Ymamoto A², Saito T², Kadoma Y² and Yu F Sasaki2,3
¹Advanced Material and Biological Engineering National Institute of Technology, Hachinohe College, Japan ²Department of Industrial System Engineering, National Institute of Technology, Hachinohe College, Japan ³Department of Pharmaceutical Health Care Faculty of Pharmaceutical Sciences, Himeji Dokkyo University, Japan
Abbreviations
ADI: Acceptable Daily Intakes; LGT: Low Melting Point.
Introduction
Food additives used in Japan are legally classified into designated additives, existing additives, natural flavors, and general food and beverage additives. Synthetic food colorings (tar dyes) are included in the designated additives, and 12 dyes are currently in use in Japan. These include 5 azo dyes, 4 xanthene dyes, 2 triphenylmethane dyes, and 1 indigo dye. Regulations on food colorings are limited to usage restrictions, and no restrictions on dosage are imposed [1]. The use of food additives at doses not exceeding the acceptable daily intakes (ADI) is generally accepted.
Among currently used food dyes, we have reported that four azo dyes, Erythrosine, Phloxisine, and Rose Bengal induced DNA damage in mouse glandular stomach, colon, and/or urinary bladder [2, 3]. These 7 dyes induced DNA damage in mouse gastrointestinal organs at a low dose (10 or 100 mg/kg). Among them, Amaranth, Allura Red, New Coccine, and Tartrazine induced DNA damage in the colon at close to the ADI [2, 3].
ADI cannot be set for food additives, pesticides and veterinary drugs when the susception of carcinogenicity by genotoxic mechanism cannot be excluded based on the absence of thresholds for genotoxicity of chemicals [4]. Nevertheless, the FDA recently revoked the authorization for the use of Erythrosine based on two studies that showed carcinogenicity in male rats by non-genotoxic mechanism: i.e., rat specific hormonal mechanism [5]. Therefore, it is serious issues whether observed genotoxicity of food dyes in mouse gastrointestinal tract (GI-tract) is related to their carcinogenicity. Despite their genotoxicity in mouse GI- tract, none of them that we showed in vivo genotoxicity has been reported to be rodent carcinogens in long-term carcinogenicity tests. To interpret the toxicological meaning of experimental genotoxicity data, it would be important the reason why there is such a large discrepancy between the results of in vivo genotoxicity and long-term carcinogenicity tests. Although carcinogenicity is examined by long-term administration in feed or drinking water, in vivo genotoxicity tests including our comet assay in mouse multiple organs have been traditionally performed by single intraperitoneal or gavage administration at MTD [2]. In our previous study, we have shown good correlation between carcinogenicity in the GI-tract and/or liver and genotoxicity increasing tendency with dosing period when carcinogens targeting the GI-tract and/or liver are given to mice continuously in the drinking water or diet [6]. Here, we examined the genotoxicity of azo and xanthene food dyes given continuously by drinking, to re-evaluate DNA damage induced by food dyes given by gavage in mouse GI-tract.
Materials and Methods
Chemicals and Animals
Tested food dyes were obtained from Tokyo Kasei Kogyo Ltd. (Tokyo, Japan). Regular (GP-42) and low melting point (LGT) agarose were obtained from Nacalai Tesque (Kyoto) and diluted, respectively, to 1% and 2% in physiological saline. Male ddY mice were obtained from Japan SLC Co. (Shizuoka, Japan) at 7 weeks of age and used after 1 week of acclimatization. They were fed commercial pellets MF (Oriental Yeast Industries Co., Tokyo, Japan) and tap water ad libitum throughout the acclimatization period and the experiment. The animal room was at 20 - 24 oC with a 12-h light-dark cycle.
The Comet Assay
The experiment followed the design of our previous comet assay studies on multiple organs [6]. Groups of 4 male ddY mice (mean body weight at starting was 37.5g) were given drinking water containing food dyes ad libitum for up to 6 days at 0.5 and 2%. The maximum concentration (2.5%) was set so that the daily intake of food dyes was not below the dose used in single gavage administration (2000 mg/kg).
Slides for comet observation were prepared after drinking for 1, 2, 4, and 6 days. Drinking started at 9:00a.m. and daily drinking water intake, body weight was measured at every 9:00a.m. During the drinking period, mice were carefully observed for pharmacotoxic signs. Mice were sacrificed at around 10:00 a.m. by bleeding under anesthesia after drinking for 1, 2, 4, and 6 days and eight organs (glandular stomach, colon, liver, kidneys, urinary bladder, lungs, brain, and bone marrow) were removed from each animal, namely, and used for the comet assay [2, 6]. When necropsies were performed, each organ was examined for changes in size, color, and texture and a small portion of each organ was fixed in 10% formaldehyde, dehydrated, and embedded in paraffin and subjected to histopathological examination.
In another experiment, each food dye was given orally to group of 4 mice by single gavage and triple gavages with the interval of 24 h. The dose for gavage was 2000 mg/ kg that is upper limit dose in the comet assay [7] and the administration volume was 10 mL/kg. From shortly after they were treated until just before they were sacrificed, mice were carefully observed for pharmacotoxic signs. Mice were sacrificed 3 and 24 hours after the final gavage, and then eight organs, (glandular stomach, colon, liver, kidney, bladder, lung, brain, and bone marrow) were removed from each individual and subjected to comet assay [2, 6]. The liver, kidney, lung, and brain were minced, suspended in 4 ml chilled homogenizing solution (pH 7.5) containing 0.075 M NaCl and 0.024 M Na2EDTA, and then homogenized gently using a Potter-Elvehjem type homogenizer at 500 - 800 rpm, in ice [2, 6].
The glandular stomach, colon, and urinary bladder were opened and rinsed with physiological saline; then the mucosa was scraped into 4 mL chilled homogenizing buffer and homogenized gently using a Potter-Elvehjem type homogenizer at 500 - 800 rpm, in ice. To obtain nuclei, the homogenate was centrifuged at 700g for 10 min at 0oC, and the precipitate was re-suspended in chilled homogenizing buffer at 1 g organ weight per milliliter [2, 6]. Seventy-five µL agarose GP-42 was quickly layered on a slide (Matsunami Glass Ind., Ltd., Osaka, Japan) coated with agarose GP-42 and covered with another slide. The slide sandwiches were placed horizontally to allow the agarose to solidify. The nucleus suspension was mixed 1:1 (v/v) with 2%, 45oC, agarose- LGT, and 75 µL of the nucleus mixture was quickly layered in the same manner after removal of the covering slide. Finally, 75 µL of agarose GP-42 was quickly layered on again. Slides prepared from nuclei isolated by homogenization were placed in a chilled lysing solution (2.5 M NaCl, 100 mM Na2EDTA, 10 mM Trizma, 1% sarkosyl, 10% DMSO, and 1% Triton X-100, pH 10) [2, 6] and kept at 0OC in the dark for about one night, then in chilled alkaline solution (300 mM NaOH and 1 mM Na2EDTA, pH 13) for 10 min in the dark at 0oC [2, 6]. Electrophoresis was conducted at 0oC in the dark for 15 min at 25 V (0.96 V/cm) and approximately 250 mA. The slides were neutralized and then stained with 50 µL of 20 µg/mL ethidium bromide (Wako Pure Chemical Industries, Ltd.) [2, 6]. We examined and photographed 50 nuclei per slide at 200 x magnification with the aid of a fluorescence microscope. The length of the whole comet (“length”) and the diameter of the head (“diameter”) were measured for 50 nuclei per organ per animal. We calculated tail length as the difference between length and diameter for each of 50 nuclei. Mean tail length of 50 nuclei from each organ was calculated for each individual animal. The differences between the averages of four treated animals and the untreated control animals were compared with the t test. A p-value less than 0.05 was considered statistically significant.
Results
The results by gavage administration and by continuous drinking were shown in Figures 1 and 2, respectively. No death, morbidity, or clinical signs were observed after any single gavage administration. Necropsy and histopathological examination of tissue sections stained by the hematoxylin- eosin revealed no treatment effect on any organ examined. Thus, any DNA damage observed was not likely to be due to general cytotoxicity (necrosis) and apoptosis. The daily intake of food dyes and body weight gain during the administration period were shown in Table 1. Daily intake of food dyes given by drinking was higher than 2000 mg/ kg. Remarkable decreases in body weight gain were not observed in the drinking period.
| Food dye | Concentration (%) | Mean daily intake | Mean | |
|---|---|---|---|---|
| Food dye (mg/kgBW) | Water (mL/mouse) | weight gain (g/mouse) | ||
| Amaranth | 0 | 0 | 6.92 | 2.3 |
| 0.5 | 859.3 | 6.15 | 2.33 | |
| 2.5 | 4555.4 | 6.54 | 1.23 | |
| Allura Red | 0 | 0 | 6.15 | 2.56 |
| 0.5 | 762.9 | 5.77 | 3.64 | |
| 2.5 | 2692.4 | 3.85 | 1.26 | |
| New Coccine | 0 | 0 | 6.15 | 3.98 |
| 0.5 | 574.6 | 4.04 | 3.51 | |
| 2.5 | 4061.3 | 6.92 | 3.14 | |
| Tartrazine | 0 | 0 | 6.92 | 2.21 |
| 0.5 | 784.4 | 6.15 | 2.6 | |
| 2.5 | 5283 | 7.69 | 1.14 | |
| Erythrosine | 0 | 0 | 5.96 | 4.69 |
| 0.5 | 706.2 | 5.38 | 4.54 | |
| 2.5 | 2362.2 | 3.65 | 1.89 | |
| Phloxisine | 0 | 0 | 5.19 | 3.91 |
| 0.5 | 739.2 | 5.38 | 4.91 | |
| 2.5 | 2256.3 | 3.85 | 3.13 | |
| Rose Bengal | 0 | 0 | 6.92 | 4.08 |
| 0.5 | 723.1 | 5.19 | 4.35 | |
| 2.5 | 2386.4 | 3.46 | 3.06 |
Table 1: Daily intake of food dyes given by drinking was higher than 2000 mg/ kg. Remarkable decreases in body weight gain were n
Azo Dyes
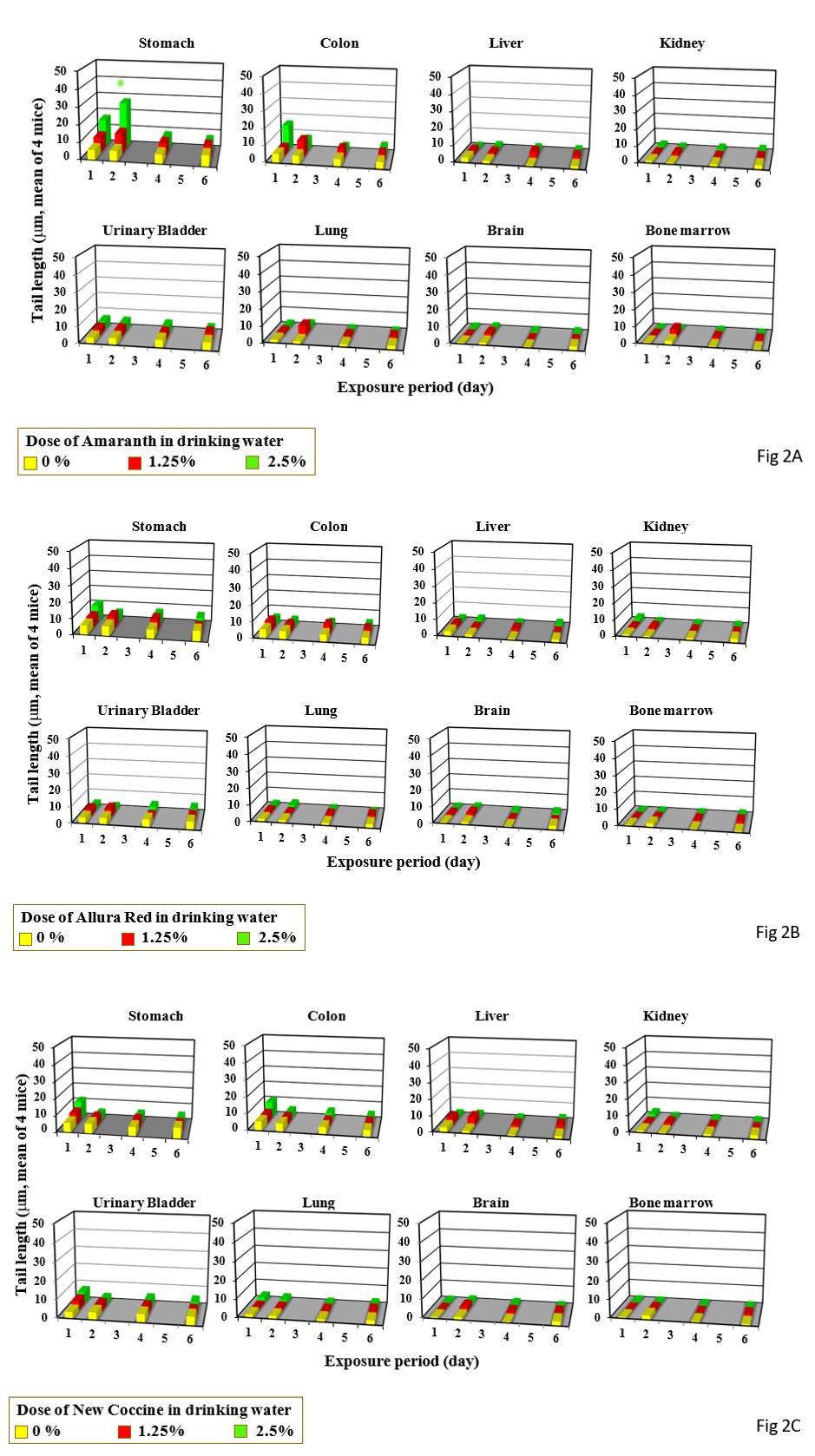
Single gavage of Amaranth at 2000 mg/kg increased tail length in the colon and urinary bladder, but its triple gavages did not increase tail length in any organs studied (Figure 1A). Drinking of Amaranth at 2.5% increased tail length only the stomach only after its 2-day drinking period (Figure 2A).
Single gavage of Arulla Red at 2000 mg/kg increased tail length in the stomach, colon, and urinary bladder, but its triple gavages at 2000 mg/kg increased tail length only in the stomach (Figure 1B). Drinkng of Arulla Red at 2.5% did not increase tail length in any organs studied the liver (Figure 2B).
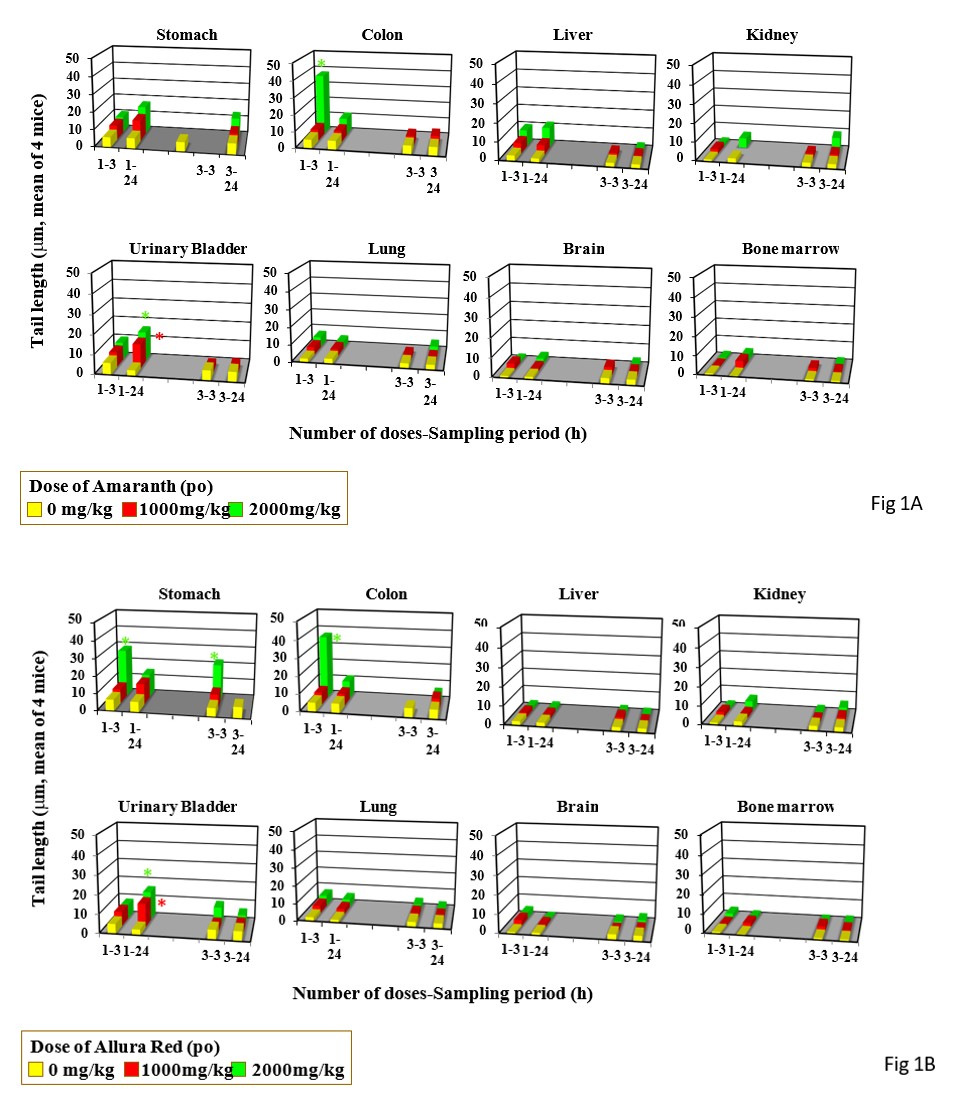
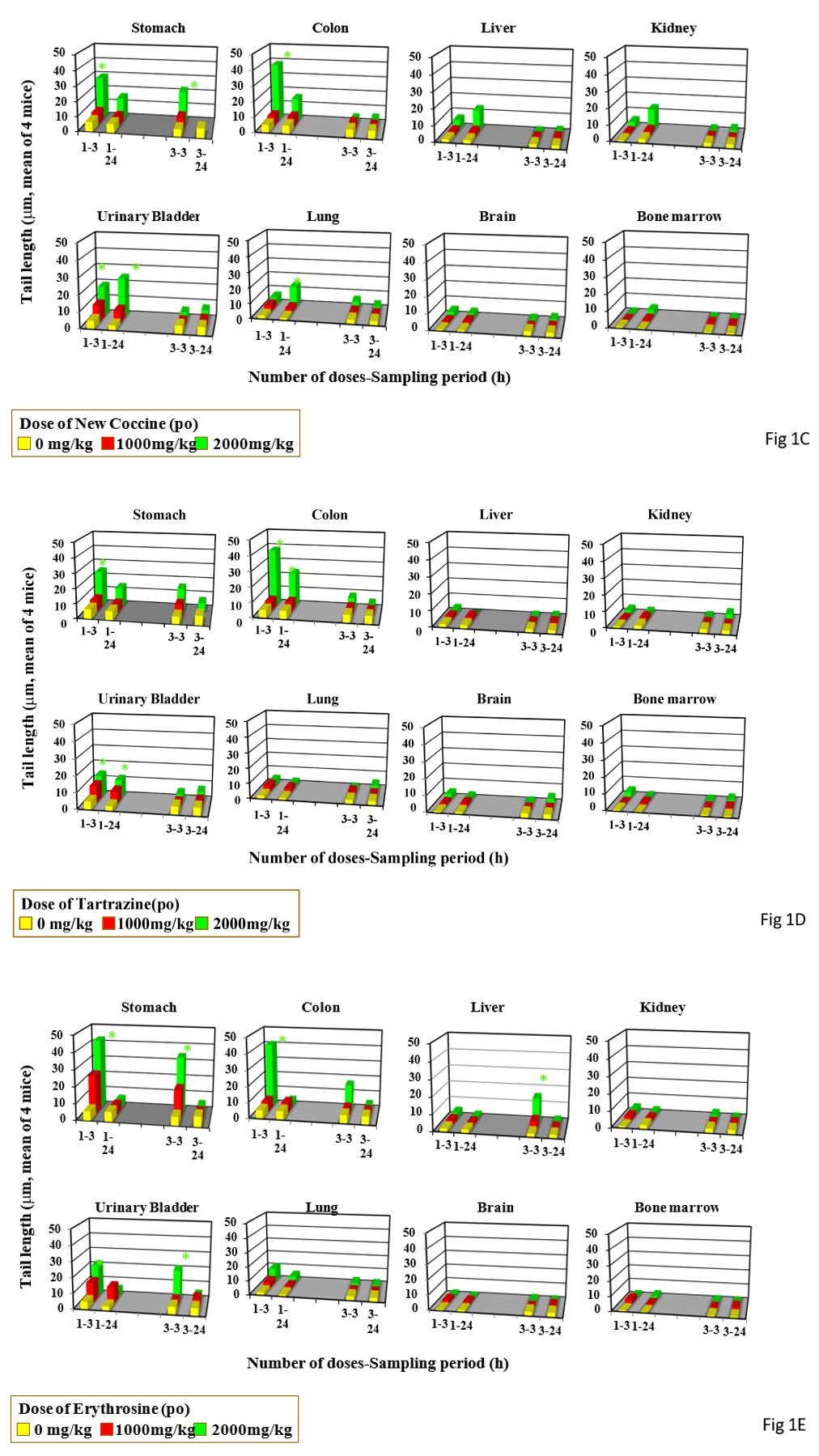
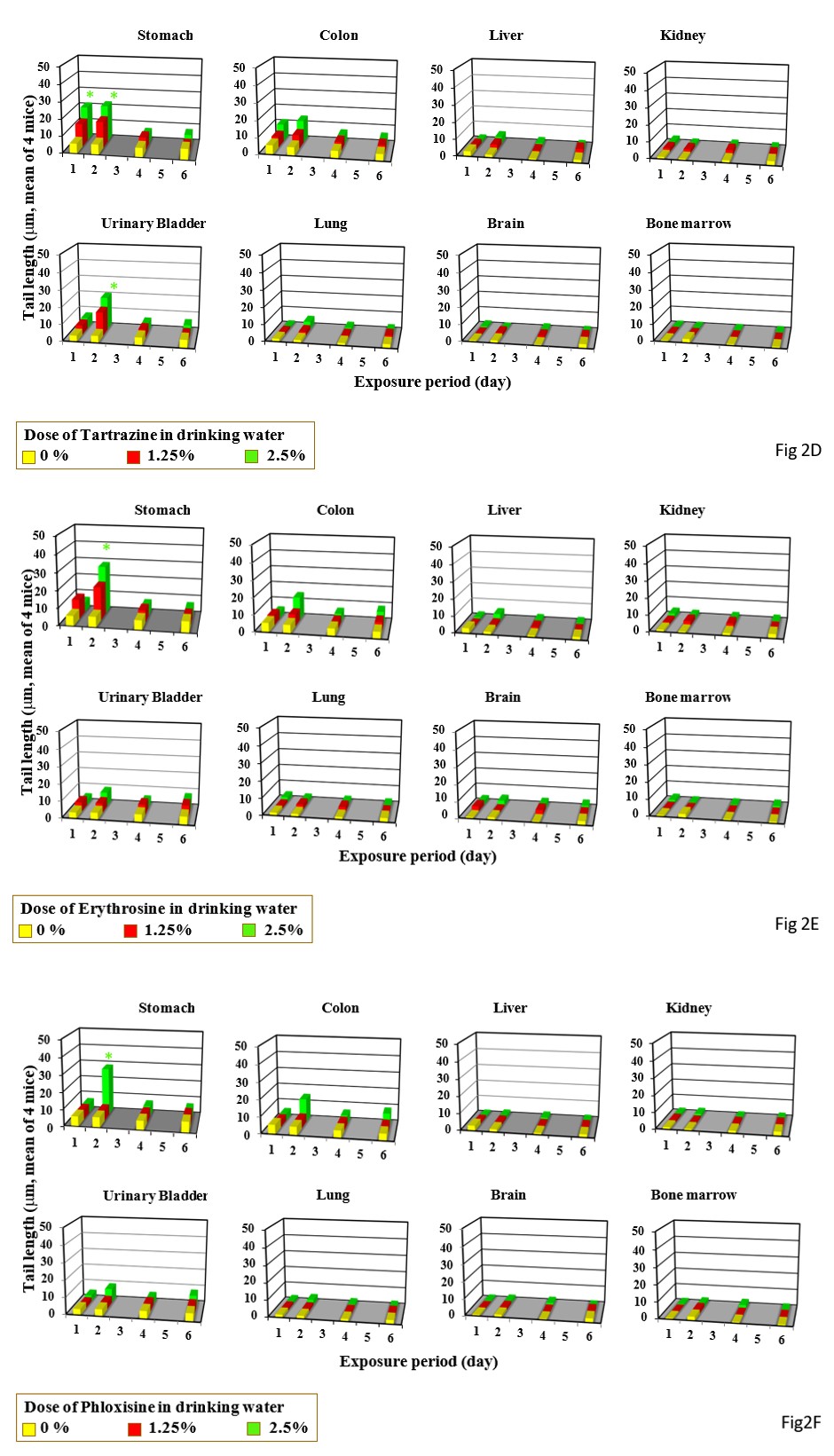
Single gavage of New Coccine at 2000 mg/kg increased tail length in the stomach, colon urinary bladder, and lung, but its triple gavages at 2000 mg/kg did not increased tail length in any organs studied (Figure 1C). Feeding of New Coccine at 2.5% mg/kg did not increase tail length in any organs studied (Figure 2C) Single gavage of Tartrazine at 2000 mg/kg) increased t in the stomach, colon and, urinary bladder. Its triple gavages at 2000 mg/kg did not increase tail length in any organs studied (Figure 1D). Drinking of Tartrazine at 2.5% increased tail length in the stomach after its 1- and 2-day drinking and in the urinary bladder h after 2-day drinking (Figure 2D).
Xanthene Dyes
Single and triple gavages of Erythrosine at 2000 mg/kg increase tail length in the stomach and colon. Its triple gavages at 2000 mg/kg increase tail length in the stomach, liver, and urinally bladder (Figure 1E). Its drinking at 2.5% increased tail length only in the stomach after its 2-day drinking (Figure 2E).
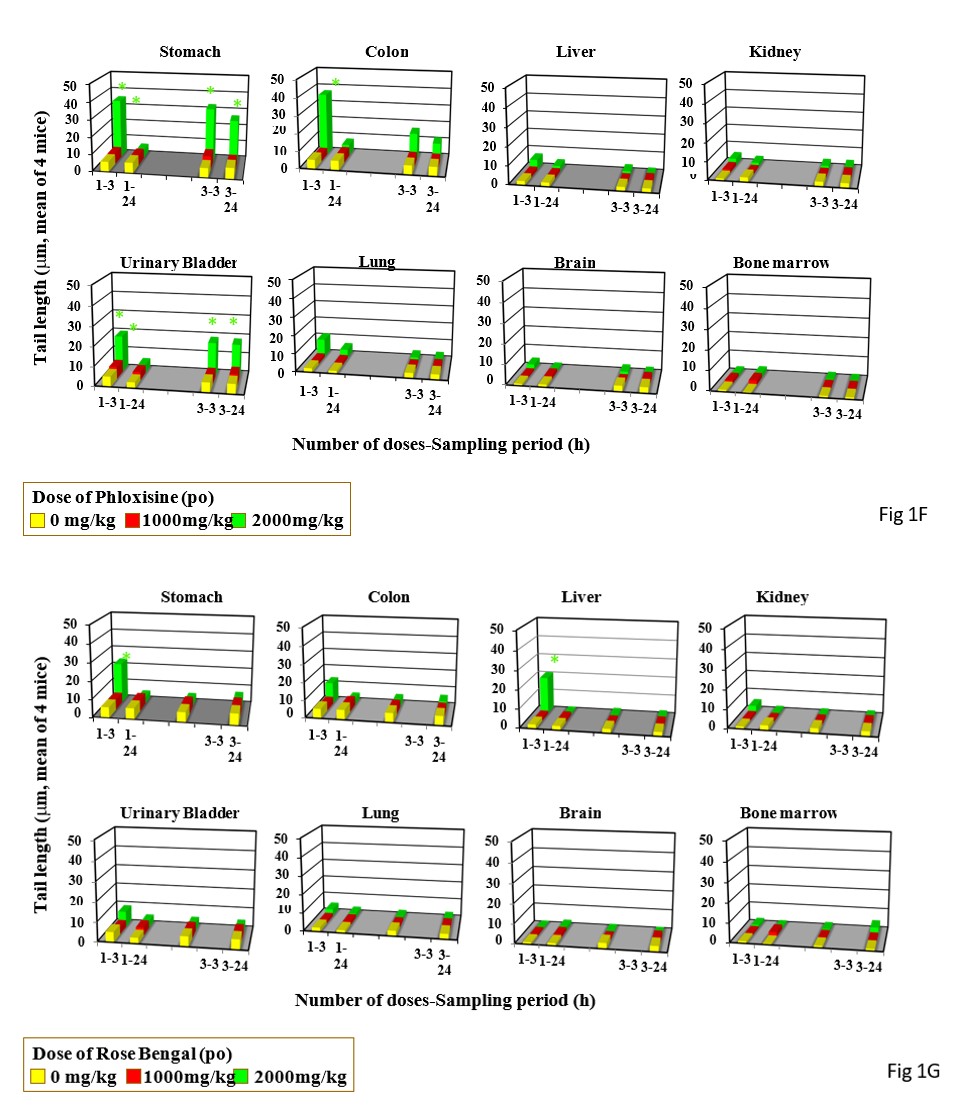
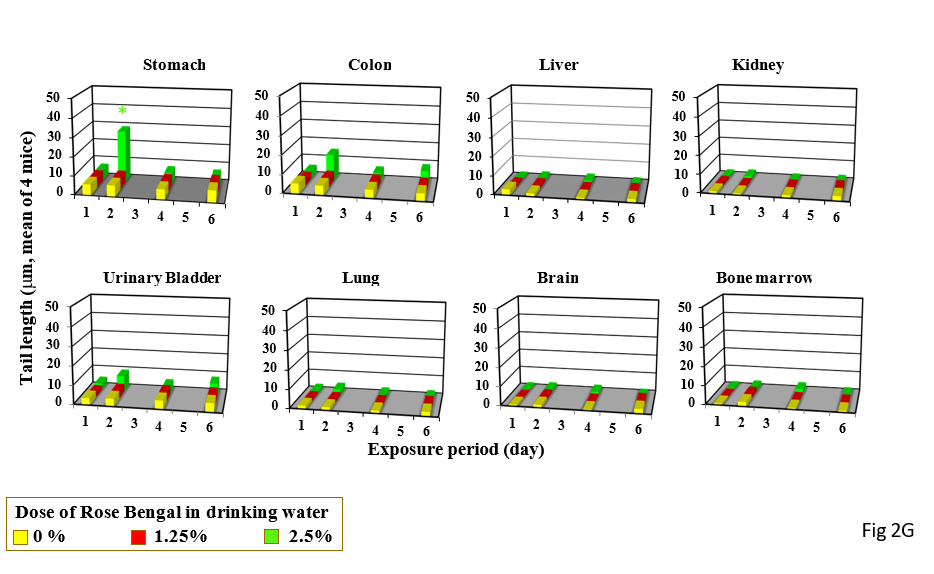
Single and triple gavages of Phloxosine at 2000 mg increase tail length in the stomach and bladders. In the colon, tail length increased by only its single gavage (Figure 1F). Its drinking at the dose used at 2.5% increased tail length in the stomach only after 2 day-drinking (Figure 2F). Single gavages of Rose Bengal at 2000mg/kg increased tail length in the stomach and liver, but its triple gavages at 2000 mg/kg did not increase tail length in any studied organs (Figure 1G). Its drinking at 2.5% increased tail length in the stomach only after 2-day drinking (Figure 2G).
Discussion
The method of the administration of food dyes greatly affected their induction of DNA damage in mouse organs. Although total intake of food dyes is 3-fold higher by triple gavages than that by single gavage, observed positive response of the comet assay was greater by single than triple gavages. Furthermore, although daily intake of food dyes by drinking at 2.5% is higher than that by single gavage at 2000 mg/kg, observed positive response of the comet assay was lower by drinking at 2.5% than single gavage at 2000 mg/kg.
In our previous study, good correlation was shown between hepatocarcinogenicity and increase of genotoxicity in the liver dependent to the duration of administration period when they were given to mice continuously in the diet or drinking water [6]. When carcinogens not targeting GI-tract were given to mice by feeding or drinking, their genotoxicity in GI-tract decreased dependently to the duration of feeding or drinking period [6]. It has been discussed that GI-tract mucosa cells having DNA damage induced in early stages of drinking or feeding might diminish due to proliferation of mucosa cells, which might prevent the accumulation of cells having DNA damage until late stage of drinking. Therefore, it could be explained the reason why DNA damage in GI-tract did not increase dependently to the duration of drinking period [6]. Although hepatocarcinogens induced DNA damage in GI-tract by gavage administration at MTD level, DNA damage in GI-tract did not increase dependently to the duration of their feeding or drinking, which coincides with the absence of their carcinogenicity in GI-tract [6].
Although food additives studied in this study induced DNA damage in GI-tract by its gavage administration at 2000 mg/kg, their drinking administration at 2.5% did not induce DNA damage in GI-tract, which would coincide with the absence of their rodent carcinogenicity [8, 9, 10, 11, 12, 13, 14, 15]. Therefore, the possibility could not be considered that their induced DNA damage by single gavage in GI-tract lead to carcinogenicity by genotoxic mechanisms. Because high dose gavage exposure to food dyes is a clinically impossible exposure pattern in humans, the results obtained from continuous exposure by drinking would be considered more toxicologically meaningful than the results from gavage administration to extrapolate observed gentoxicity to humans [16]. Gavage is appropriate for testing orally administered drugs, although none of the food dyes studied is used as a drug [17]. The results of this study might indicate the difficulty to extrapolate directly the positive genotoxicity results from acute administration to humans.
Abe, et al. [18] further studied genotoxicity of Amaranth in mouse colon and observed followings; (1) single intraperitoneal injection of Amaranth at 1000 mg/kg or higher induced DNA damage in mouse colon, (2) single gavage of Amaranth did not yield DNA damage in mouse colon received choledocholigation, from which it has been discussed that intestinal bacterial metabolites of Amaranth were absorbed from the intestinal tract and returned to the intestinal tract via enterohepatic circulation, inducing DNA damage in GI-tract [18]. Azo dyes are reduced to aromatic amines in the GI-tract by azo reductase synthesized in the gastrointestinal wall cells and microflora, from which it has been discussed that reduced products of Amaranth are further activated metabolically in the liver and yielded metabolites returned to the GI-tract by enterohepatic circulation to show its genotoxicity in mouse colon [18]. Triple gavages of azo food dyes led to negative response in mouse GI-tract. The possible explanation for this elimination might be that their triple gavages lower the activity of azo reduction in the GI-tract and/or the sensitivity of GI-tract mucosa cells against azo metabolites as we have previously discussed [6]. p-Dimethylaminoazobenzene yielded DNA damage in the liver after triple and single gavage, which might suggest that azo reduction occurred even after triple gavages [6]. Amaranth caused DNA damage mainly in the GI- tract in mice but not in rats, suggesting that there are species differences in the metabolism of intestinal bacteria playing an important role to show genotoxicity of Azo dyes [18].
Therefore, observed genotoxicty of azo dyes could not be directly extrapolated to human without the consideration about azo reduction in human. The results from continuous administration by drinking or feeding are likely to reflect carcinogenicity better than the results by single gavage administration as discussed above. Therefore, despite the presence of positive responses in GI-tract by single gavage of food dyes, the absence of positive response in mouse GI-tract by continuous drinking administration of food dyes could be considered to show their absence of carcinogenicity by genotoxic mechanisms.
Acknowledgements
This research was conducted in Advanced Material and Biological Engineering Course, National Institute of Technology, Hachinohe College as part of a graduation research project by Natsue Yokohama under the research guidance of course staffs based on allocation of school educational expenses. The authors acknowledge the Advanced Material and Biological Engineering Course, National Institute of Technology, Hachinohe College. References
1. MHW (1959) Specifications and standards for food additives.
2. Sasaki YF, Kawaguchi S, Kamaya A, Ohshita M, Kabasawa K, et al. (2002) The comet assay with eight mouse organs: Results with 39 currently used food additives. Mutation Res 519: 103-119.
3. Tsuda S, Murakami M, Matsusaka N, Kano K, Taniguchi K, et al. (2001) DNA Damage Induced by Red Food Dyes Orally Administered to Pregnant and Male Mice. Toxicol Sci 61: 92-99.
4. Nhomi T (2008) Possible mechanisms of practical Thresholds for Genotoxicity. Genes and Environment 30(4): 108-113.
5. FDA (2025) To revoke authorization for the use of red No. 3 in food and ingested drugs.
6. Yokohama N, Kawaguchi S, Yamamoto A, Nakamura T, Sasaki YF (2025) Study of genotoxicity of hepatocarcinogens in multiple organs in mice by feeding and drinking using the comet assay. Adv Clin Toxicol: 16000327.
7. OECD (2016) Test No.489: in vivo mammalian alkaline comet assay.
8. Borzelleca JF, Hallagan JB (1988) A chronic toxicity/ carcinogenicity study of FD & C Yellow No. 5 (tartrazine) in mice. Food and Chem Toxicol 26: 189-194.
9. Borzelleca JF Hallagan JB (1988) Chronic toxicity/ carcinogenicity studies of FD & C Yellow No. 5 (tartrazine) in rats. Food and Chem Toxicol 25: 179-187.
10. Borzelleca JF, Olson JW, Reno FE (1989) Lifetime toxicity/ carcinogenicity study of FD & C Red No. 40 (Allura Red) in Sprague-Dawley rats. Food Chem Toxicol 27: 701-705.
11. Borzelleca JF, Olson JW, Reno FE (1991) Lifetime toxicity/ carcinogenicity studies of FD & C red No. 40 (Allura Red) in mice. Fd Chem Toxicol 29: 313-319.
12. Maekawa A, Matsuoka C, Onodera H, Tanigawa H, Furuta K, et al. (1987) Lack of carcinogenicity of tartrazine (FD & C Yellow No. 5) in the F344 rat. Fd Chem Toxicol 25: 891-896.
13. Borzelleca JF, Hallagan JB (1987) Lifetime toxicity/ carcinogenicity study of FD & C Red No. 3 (erythrosine) in mice. Food & Chemical Toxicol 25: 735-737.
14. IARC (1975) IARC Momographs on the Evaluation of the carcinogenic Risk of Chemicals to Humans, Some Aromatic Azo Compounds. International Agency for Research on Cancer 8: 41-51.
15. IARC (1987) Overall evaluation of carcinogenicity, An updating of IARC Monographs. Vol 1-42, (Suppl 7): 56.
16. Weil CS (1972) Guideline for experiments to predict the degree of safety of a material for man. Toxicol Appl Pharmacol 21: 194-199.
17. Johanso FM (2002) How many food additives are rodent carcinogens?. Environ Mol Mutagenesis 39: 69-80.
18. Abe T, Shimada C, Satoh I, Tsuda S, Sasaki YF (2002) Differential Effects of Red Food Dies on Colon DNA Damage between Rats and Mice. Annual Meeting of Japanese Veterinary Science Society.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells
- The Role of Faculty in Assessment: Traditional vs. Competency- Based Medical Education in Toxicology Learning