Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
Macrophages are key players in innate immunity and inflammation during viral infections, including COVID-19. This study explored the cellular and proteomic responses of THP-1-derived macrophages following exposure to SARS-CoV-2. Despite lacking ACE2 expression, these cells internalized the virus, although no productive replication was observed up to 48 hours post-infection. The virus triggered an inflammatory response marked by increased secretion of TNF-α, IL-6 and IL-10 at 24 hours, and elevated levels of TNF-α, IL-6 and GM-CSF at 48 hours post-infection. Proteomic analysis revealed early downregulation of G6PD and upregulation of FABP4 at 24 hours post-infection, suggesting metabolic reprogramming. At 48 hours, a broader set of proteins showed reduced abundance, including those involved in NADH metabolism, cytokine mediated signaling and endoplasmic reticulum (ER) stress pathways. These proteomic insights, while requiring molecular validation, macrophage responses in COVID-19 pathogenesis, highlighting the THP-1 model's value for investigating host inflammatory and stress responses independent of productive viral replication, and revealing key proteins and pathways involved in COVID-19 pathogenesis.
Abbreviations
ER: Endoplasmic Reticulum; WHO: World Health Organization; ARDS: Acute Respiratory Distress Syndrome; G6PD: Glucose-6-Phosphate Dehydrogenase; FABP4: Fatty Acid-Binding Protein 4; MOI: Multiplicity of Infection; LVCM: Laboratory of Clinical and Molecular Virology; PBS: Phosphate-Buffered Saline; DTT: Dithiothreitol; FASP: Filter-Aided Sample Preparation; LFQ: Label-Free Quantification; MDM: Monocytes-Derived Macrophages; NADPH: Nicotinamide Adenine Dinucleotide Phosphate; ER: Endoplasmic Reticulum; UPR: Unfolded Protein Response.
Introduction
Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2), the etiological agent of Coronavirus disease 2019 (COVID-19), has caused a global health crisis since its emergence. By May 2025, the World Health Organization (WHO) had documented over 700 million cases of COVID-19 globally [1]. Despite significant advancements in vaccines and therapies development, COVID19 has underscored the critical role of the innate immune system in responding to viral infections, both in pathogen clearance and in regulating inflammatory responses [2, 3].
Macrophages are versatile components of the immune system that differentiate into distinct functional states in response to environmental stimuli. The M1 phenotype is typically associated with antiviral and pro-inflammatory roles, marked by the secretion of cytokines such as IL- 6, TNF-α, and type I interferons, all contributing to effective immune clearance. In COVID-19, dysregulation in macrophage activation is linked to the development of cytokine storms and tissue damage. In contrast, alternatively activated macrophages (M2) are associated with anti- inflammatory functions, tissue repair, and immunoregulation through the secretion of IL-10 and TGF-β [4, 5]. Previous studies suggest that although SARS-CoV-2 primarily infects epithelial cells via receptors such as ACE2 and TMPRSS2 [6], the susceptibility of human macrophages to the virus also dependent on these receptors’ expression, however they do not sustain productive virus replication [7].
Moreover, activation of macrophages upon SARS-CoV-2 exposure and uptake has been associated with the cytokine storm observed in severe cases of COVID-19, contributing to tissue damage and worsening clinical outcomes [8, 9]. SARS-CoV-2 infection promoting dysregulation of the M1/ M2 balance has been observed and is thought to contribute to disease [10]. Viral proteins have been shown to interfere with macrophage activation, promoting immune evasion by suppressing type 1 interferon responses [11], however Increasing an excessive activation of M1 macrophages witch has been implicated in the cytokine storm and acute respiratory distress syndrome (ARDS), highlighting the complex role of macrophage polarization in COVID-19 pathophysiology [4].
Comprehensive molecular studies using omics-based approaches have helped respond to how SARS-CoV-2 infection modulates cellular processes and molecular consequences of viral entry, replication, and tropism, as well as the downstream effects on host disease progression [12, 13]. Compared to studies based on isolated viral proteins or recombinant constructs, analyses using the native virus provide a more physiologically relevant model for understanding the full spectrum of viral-host interactions. In this study, we assessed the inflammatory and proteomic responses of THP-1-derived macrophages following exposure to SARS-CoV-2. Although the infection was non-productive, viral particles were rapidly internalized upon contact. This abortive interaction nevertheless provoked a strong immune activation, evidenced by increased levels of TNF-α, IL-6, and IL-10 at 24 hours, and heightened expression of TNF-α, IL-6, and GM-CSF at 48 hours post-exposure. Additionally, proteomic profiling demonstrated time-dependent alterations in macrophage metabolic and immune signaling pathways. Early post- infection changes included upregulation of glucose-6- phosphate dehydrogenase (G6PD) and fatty acid-binding protein 4 (FABP4), indicative of shifts in redox balance and lipid metabolism. At later stages, there was a marked reduction in the abundance of proteins associated with NADH-dependent processes, cytokine signaling pathways, and markers of endoplasmic reticulum (ER) stress [14, 15].
Material and Methods
SARS-CoV-2 Stock Production
The SARS-CoV-2/SP02/2020/BRA strain (GenBank accession MT126808.1), supplied by the Laboratory of Clinical and Molecular Virology (LVCM) at ICB-USP under the national research network of Brazil [16] was propagated under biosafety level 3 (BSL-3) conditions in accordance with international biosafety guidelines. Viral amplification was conducted in Vero CCL-81 cells (ATCC) maintained in VP-SFM medium (Thermo Fisher) within T225 culture flasks. Infections were initiated at a multiplicity of infection (MOI) of 0.05, and supernatants were collected after 72 hours post-infection (hpi). The collected medium was centrifuged at 3,000 × g for 10 minutes to remove cell debris, and the clarified viral suspension was stabilized in SPG buffer and stored at –80°C. Viral titers were determined by tissue culture infectious dose (TCID50) assay [17, 18] using Vero cell monolayers. For mock infections, uninfected Vero cells were handled under identical conditions, and the resulting supernatants were processed similarly to ensure matched experimental controls.
THP-1 Cell Maintenance and Differentiation
THP-1 human monocytic cells (ATCC® TIB-202™) were cultured in RPMI-1640 medium (Sigma R6504) supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY, USA, #26140079), 2 mM L-glutamine (Sigma- Aldrich St. Louis, MO, USA, #A2916801), and 1 mM sodium pyruvate (Sigma-Aldrich, Sigma-Aldrich, St. Louis, MO, USA, #P5280). Cells were incubated at 37°C in a 5% CO₂ humidified environment and passaged every 48–72 hours. To differentiate monocytes into macrophage-like cells, 25 nM of phorbol 12-myristate 13-acetate (Sigma-Aldrich, St. Louis, MO, USA, #P8139) was added for 48 hours. Following this exposure, cells were incubated in fresh PMA-free medium for an additional 24 hours to allow adherence and phenotypic maturation, as previously described in in vitro inflammation models [19].
SARS-CoV-2 Infection of Macrophages
Differentiated macrophages seeded in 6-well plates (8×10⁵ cells/well) were exposed to SARS-CoV-2 at an MOI of 1. Viral adsorption was performed over 60 minutes at 37°C. Following this step, the inoculum was removed and replaced with fresh RPMI complete medium. Infected cells were incubated for 24 and 48 hours, and mock- infected macrophages, treated with control supernatants, were maintained under the same conditions to serve as experimental controls.
Kinetics of Infection and Viral Titration
To evaluate infection kinetics, cell cultures were monitored at 24 and 48 hpi. Supernatants were collected at both time points and subjected to viral titration. Serial ten- fold dilutions of each sample were prepared and applied to Vero cell monolayers cultured in multi-well plates. After 72 hours, cytopathic effects were assessed by light microscopy. Infectious titers were calculated using the Reed-Muench method [18].
Cytokine and Chemokine Analysis
Supernatants from infected and control macrophages were analyzed for cytokine and chemokine production using a multiplex magnetic bead-based immunoassay (Millipore HCYTOMAG-60K-13). This panel detects nine inflammatory mediators: TNF-α, IFN-α, IL-1β, IL-6, IL-4, IL-10, GM-CSF, CXCL10, and CCL2. Measurements were carried out using the Luminex-200 system operated via xPONENT 4.3 software. Data were processed using MILLIPLEX Analyst 5.1 software, applying manufacturer-specified calibration curves and sensitivity thresholds.
Sample Preparation and LC–MS/MS Proteomic Analysis
At 24 and 48 hpi, macrophage supernatants were removed, and cells were washed three times with cold phosphate-buffered saline (PBS). Harvested cells were resuspended in ice-cold lysis buffer containing 2 M urea and 5 mM dithiothreitol (DTT), followed by storage at –80°C. For protein extraction, samples were thawed on ice and subjected to ultrasonication (five cycles of 30 seconds on/ off). Lysates were clarified by centrifugation (15,000 × g, 30 minutes, 4°C), and proteins were digested using the filter- aided sample preparation (FASP) method with 10 kDa cut-off filters (Merck Millipore, Darmstadt, Hessen, DE, USA), Tryptic digestion occurred overnight at 37°C. Peptides were purified using StageTips packed with SDB-XC membranes, vacuum- dried, and reconstituted in 0.1% formic acid. For LC–MS/MS analysis, 250 ng of each sample was injected into a nano-LC EASY 1200 (LC-030378, Thermo Fisher Scientific, Waltham, MA, USA) system coupled with a Q Exactive Plus (03893L, Thermo Fisher Scientific, Waltham, MA, USA) spectrometer at the Mass Spectrometry Unit of CENTD (Butantan Institute, Butantã, Brazil). Chromatographic separation was achieved using a PepMap100 C18 trap column (75 μm × 20 mm, 3 μm) in line with a PepMap RSLC analytical column (50 μm × 150 mm, 2 μm) at a flow rate of 200 nL/min. The mobile phases - solvent A (0.1% v/v formic acid) and solvent B (80% v/v acetonitrile containing 0.1% v/v formic acid) - were used in a linear gradient (5–30% B in 50 min, 30–60% B in 13 min, 60–100% B in 2 min, and a step of 100% B for 5 min). Mass spectrometry was performed in positive ion mode across a 300–1500 m/z range, selecting the top seven precursors for higher energy collisional dissociation (HCD) fragmentation. Parameters included AGC targets of 3×10⁶ (full scans) and 2×10⁵ (MS/MS), maximum injection times of 200 ms (full) and 120 ms (MS/MS), and a dynamic exclusion of 60 seconds.
Proteomics Data Processing
Mass spectrometry raw data were analyzed using the MaxQuant software platform. Protein identification was carried out via the Andromeda search engine, referencing the Homo sapiens UniProt database (retrieved in May 2022). Precursor and fragment ion mass tolerances were set at 4.5 ppm and 20 ppm, respectively. Carbamidomethylation of cysteine residues was applied as a fixed modification, whereas oxidation of methionine and N-terminal acetylation were included as variable modifications. Tryptic digestion was specified, allowing up to two missed cleavage sites per peptide. Protein and peptide identifications were both constrained to a maximum false discovery rate (FDR) of 1%. For protein-level FDR estimation, a decoy database composed of reversed protein sequences was utilized. Only proteins identified with a minimum of two peptides (unique and razor) were retained. Quantification was performed using label-free quantification (LFQ) based on normalized intensity values generated by MaxQuant. Default parameters were maintained unless stated otherwise. Prior to statistical evaluation, data were logarithmically transformed, and entries flagged as contaminants, reverse hits, or identified solely by site modifications were excluded, along with any features missing across replicates.
Functional Network Analysis
To explore biological functions, protein interaction data were analyzed using Cytoscape (version 6) with the ClueGO (v4) and AutoAnnotate (v5) plugins. ClueGO constructs functionally grouped networks based on gene ontology (GO) biological process enrichment, displaying nodes as terms and edges according to kappa statistics reflecting shared genes. A hypergeometric test was applied for enrichment assessment, with Bonferroni step-down correction used to adjust for multiple comparisons. Only terms with corrected p-values below 0.05 were visualized. Semantic grouping of related GO terms was facilitated using AutoAnnotate to enhance interpretability of the networks.
Statistical Analysis
All experiments were performed in three independent biological replicates, each analyzed in duplicate. Quantitative data for viral titers and multiplex assays are presented as mean ± standard deviation (SD). Group comparisons between SARS-CoV-2-infected and mock-treated conditions were made using non-parametric Student’s t-tests. These tests were conducted using GraphPad Prism 6.01 (GraphPad Software), with statistical significance defined as p < 0.05. For proteomic comparisons, either two-tailed Student’s t-tests or one-way ANOVA (for multigroup comparisons) were applied using the Perseus software suite (version 3), incorporating permutation-based FDR control at 5% with an S₀ threshold set to 0.1. Visual representations of statistical outputs were generated in R using standard packages including ggplot2 and pheatmap. The number of replicates per experiment is detailed in the respective figure legends.
Results
THP-1 macrophage susceptibility to SARS-CoV-2
Viral titration assay was performed to evaluate the TCID50 (Tissue culture infectious dose 50%) of macrophages using MOI 1. The results showed that after 1 hour of adsorption, there was internalization of ≅ 2x102 viral particles. The macrophages presented titers of 2.3x103 TCID50/ml at 24 hpi and 3.7 x102 TCID50/ml at 48 hpi, indicating non-productive infection (Figure 1). Macrophages are specialized phagocytic cells and can uptake virus particles independent of specific receptor interaction. Possibly, the SARS-CoV-2 uptake route in these cells occurs independently of the ACE2, TMPRSS2, and CD147 receptors, since there was no expression of these receptors by RT-qPCR (data not showed), consistent with previous reports that THP1 do not express ACE2 [7].
Macrophage Cytokine and chemokine release induced by SARS-Cov-2
To evaluate the inflammatory effect of SARS-CoV-2 exposure on macrophages, the release of cytokines and chemokines was investigated by multiplex 24 and 48 hpi. SARS-Cov-2 induced release of IL-6 (17.01 ± 3.325 and 3.793 ± 0.64 pg/mL for Sars-CoV-2 and mock, respectively, p = 0.0175), CCL2 (727.5 ± 82.13 and 380.2 ± 24.37 pg/mL for Sars-CoV-2 and mock, respectively, *p = 0.0154), and IL- 10 (302.4 ± 42.33 and 132.3 ± 2.36 pg/mL for Sars-CoV-2 and mock, respectively, p = 0.0160) after 24 hpi Figure 2. Subsequently, there was an increase in the release of TNF-α (1263 ± 130.8 and 539.4 ± 93.22 pg/mL for Sars-CoV-2 and mock, respectively, p = 0.0108) IL-6 (29.18 ± 6.40 and 3.44 ± 0.26 for Sars-CoV-2 and mock, respectively p = 0.0158), and GM-CSF (178.2 ± 14.71 and 110.0 ± 6.61, for Sars-CoV-2 and mock, respectively, *p= 0.0134) after 48 hpi. There was no significant modulation of the other cytokines tested (IL-1β, IL-4 and CXCL10). We also evaluated the release of IFNα2, however, all samples were below the method’s detection limit.
Proteomic Analysis
To investigate changes in the proteome of macrophages, cells were exposed to SARS-CoV-2 or mock for 1 h and incubated for 24 and 48 hpi. Across all replicates, 1248 and 577 protein groups were quantified in macrophage samples after 24 and 48 hpi, respectively. After statistical analyses proteins with p <0.05 were considered differentially abundant. The results identified 3 differentially abundant (DA) proteins at 24 hpi and 71 DA proteins at 48 hpi Figure 3.
Among the group of DA proteins at 24 hpi, Glycine-- tRNA ligase (GARS1) (FC= -2.31) and Glucose-6-phosphate dehydrogenase (G6PD) (F = -2.26) were reduced after infection with SARS-CoV-2 and Fatty Acid-Binding Protein 4 (FABP4) (FC= 6.15) was increased.
At 48 hpi, DA proteins showed reduced abundance in 71 proteins. Enrichment analysis using the ClueGO and AutoAnnotate applications available in the Cytoscape software showed a prominent cluster of proteins associated with pathways related to the Protein stabilization and telomerase maintenance, NADH metabolic process, and response to protein unfolding. In relation to the inflammatory response, the enriched pathways indicate correlation with the Response to IL-7, IL-12 mediated signaling pathway and IL-4 response. Among the downregulated proteins the top ten were Profilin-1 (PFN1) (-2.29), Tropomyosin alpha-4 chain (TPM4) (-2.29), Gelsolin (GSN) (-2.15), Cathepsin G Macrophages Titers:MOI- 1
8 - - - - - - - - - - - - - - - - - - - - - - - - - - -
7
Infections titers TCID50/mL Log10 scale
1 24 48 0 Figure 1: Lack of productive SARS-CoV-2 infection in THP-1 macrophages. Cells were either infected with SARS-CoV-2 at a multiplicity of infection (MOI) of 1 or mock-treated for one hour, and subsequently maintained in culture for 24 and 48 hours. Data shows viral titer measurement by TCID50 assay of THP-1 macrophages. Values shown are the means ± standard deviation (SD).
(CTSG) (-2.10), Endoplasmin (HSP90B1) (-2.00), Enhancer of rudimentary homolog (ERH) (-1.77), F-actin-capping protein subunit alpha-1 (CAPZA1) (-1.75), Plastin-2 (LCP1) (-1.69), 6-phosphogluconate dehydrogenase (PGD), and Vimentin (VIM) (-1.44).
Inoculum Titer
(hpi)
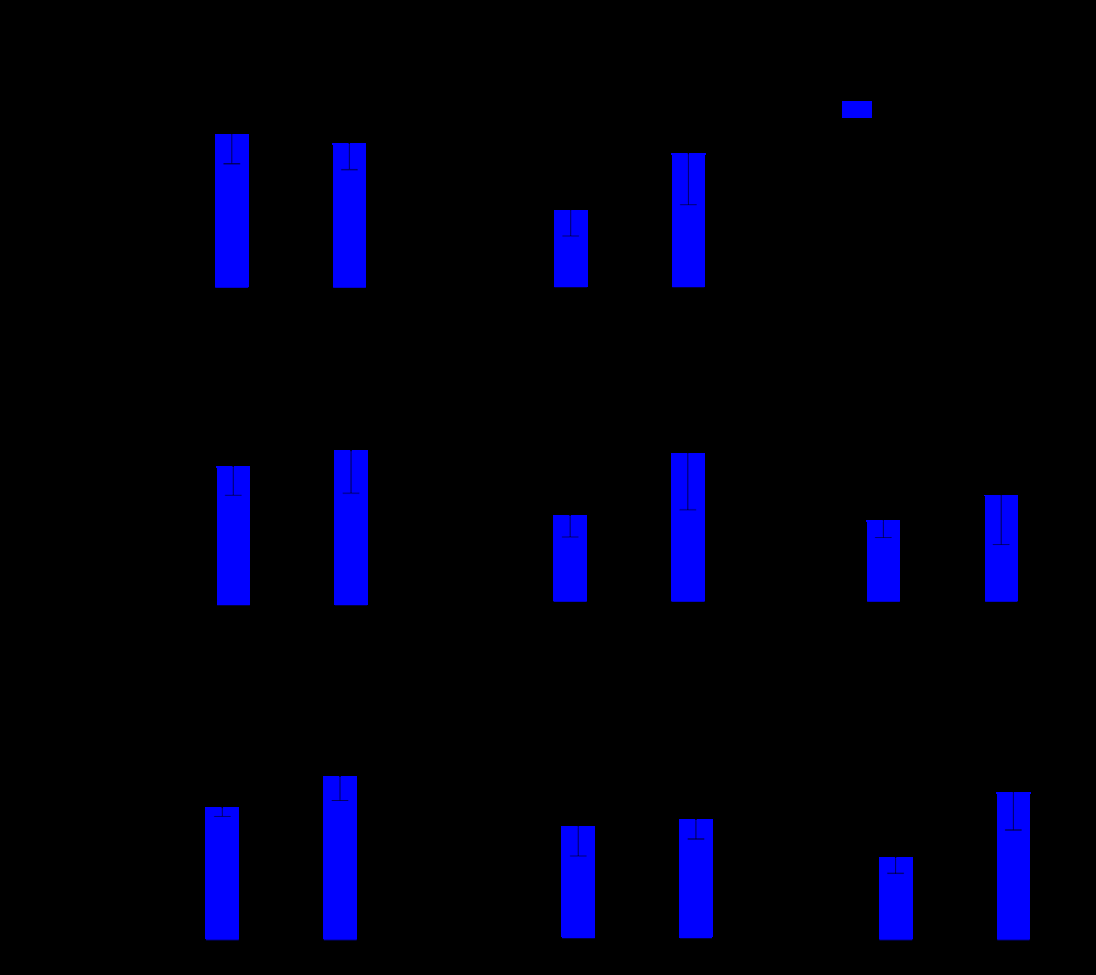
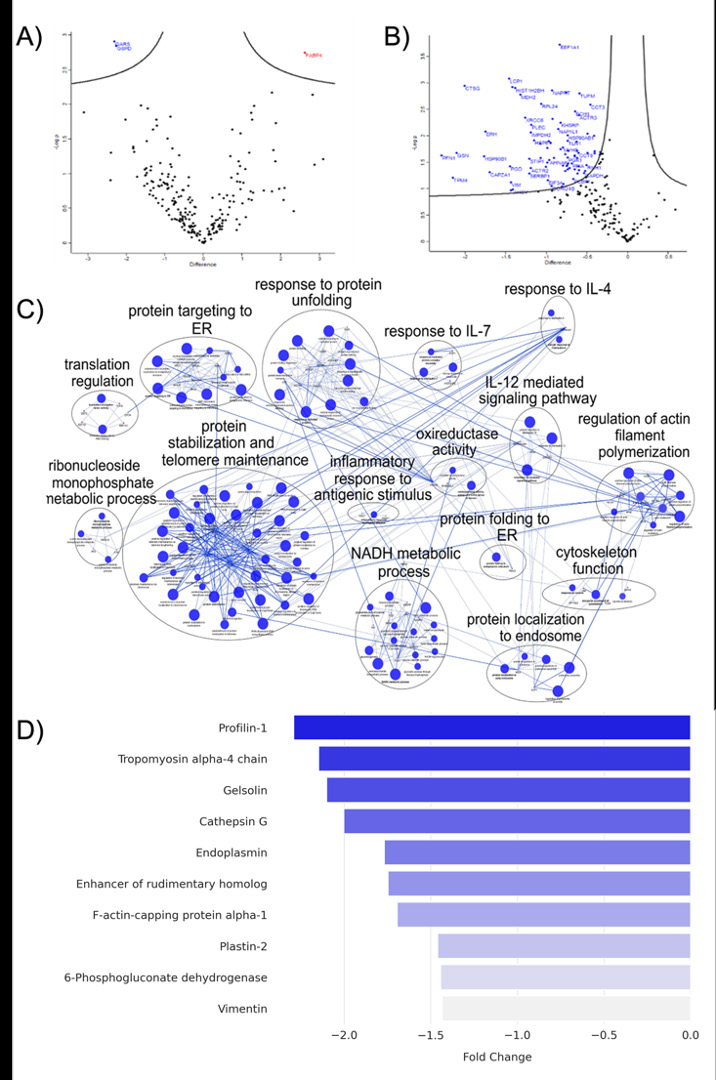
Figure 3: Proteomic and Functional Enrichment Analysis of SARS-CoV-2-Exposed THP-1 Macrophages. (A e B) Volcano plots highlight significantly high-abundance (red) and low-abundance (blue) protein groups after 24 e 48 hpi. (C) ClueGO network visualization of enriched pathways among low-abundance proteins at 48 hpi. (D) Top 10 low-abundance proteins after 48hpi.
Discussion
Infection with SARS-CoV-2 is associated with an intensified activation of bronchoalveolar immune cells, resulting in sustained overproduction of pro-inflammatory cytokines, including IL-6, TNF-α, and IL-1β [20, 21, 22]. Severe cases are further characterized by a breakdown in the normal resolution of inflammation, evidenced by the buildup of inflammatory macrophages within the alveolar spaces and ongoing secretion of chemokines and fibrotic signaling molecules. Although substantial progress has been made, the exact pathways through which SARS-CoV-2 disrupts immune homeostasis remain insufficiently defined. Critical aspects such as the virus’s interaction with macrophages, the resulting immune dysregulation, cellular impairments, and their roles in immune escape and viral spread are still under active exploration. In this investigation, we analyzed cytokine secretion dynamics and the proteomic landscape of macrophages derived from the THP-1 cell line following 24- and 48-hour exposures to the original Wuhan variant of SARS-CoV-2.
Given that macrophages are highly efficient phagocytic cells, their susceptibility to SARS-CoV-2 infection has been widely discussed. SARS‐CoV‐2 can enter primary monocytes and monocytes-derived macrophages (MDM). However, there is consensus about the inability of SARS‐ CoV‐2 to replicate in these cells in relation to poor expression of SARS‐ CoV‐2 receptors, ACE2 and TMPRSS2 [23, 24, 25, 26, 27].
Viral particles can enter these cells through mechanisms such as endocytosis and phagocytosis, as well as via ACE2- mediated entry in populations that express the receptor. Although THP-1 monocytes and their derived macrophages do not express ACE2, they have also been reported to permit viral entry, but do not support productive replication. Recent study demonstrated that SARS-CoV-2 can replicate and generate infectious particles in THP-1 cells engineered to overexpress ACE2, indicating dependence on the receptor expression [7]. Our findings support the non-permissiveness of the THP-1-derived macrophage lineage to SARS-CoV-2 replication. Once viral uptake was observed within 1-hour post-infection, no increase in viral titer was detected at 24 and 48 hpi, indicating the absence of productive infection. Despite the absence of productive replication, macrophages are highly responsive to viral antigens, initiating robust immune activation [28, 29, 30, 31]. Our results reveal that macrophages respond to SARS-CoV-2 exposure and internalization with a marked pro-inflammatory reaction, as indicated by increased secretion of cytokines such as TNF-α and IL-6 at both 24- and 48-hours post- infection. This inflammatory signaling has been linked to the activity of various structural and accessory proteins of the virus. Among these, the Envelope (E) protein of SARS-CoV-2 has been specifically implicated in triggering pyroptosis and amplifying the production of inflammatory mediators— including TNF-α, IL-6, and IL-1β—in THP-1 cells by engaging key signaling pathways such as NF-κB, JNK, and p38 [32]. Similarly, the Nucleocapsid (N) protein can trigger substantial cytokine release, contributing to systemic inflammation [30, 31, 32, 33, 34]. Furthermore, the direct effect of the SARS-CoV-2 spike protein on IL-6 release has been previously described, highlighting its role in activating the STAT3 transcription factor pathway [35, 36].
To further elucidate the molecular pathways underlying these effects, we conducted a proteomic analysis of macrophages infected with SARS-CoV-2. This analysis identified Fatty Acid Binding Protein 4 (FABP4) as the most highly expressed protein in THP-1-derived macrophages 24 hours post-infection. FABP4 has been identified as a critical host factor actively involved in facilitating SARS- CoV-2 replication, specifically by being recruited to and influencing the structural integrity and organization of viral replication organelles (ROs) [37, 38, 39]. Previous studies have demonstrated that elevated FABP4 levels in patients and FABP4+ alveolar macrophages are dominant in mild to moderate COVID‐19 patients [40].
Conversely, results showed glucose-6-phosphate dehydrogenase (G6PD) with low-abundance after 24 hpi. G6PD serves as a key regulatory enzyme in the pentose phosphate pathway (PPP), playing a critical role in the production of nicotinamide adenine dinucleotide phosphate (NADPH) [41]. Proteomic profile of THP-1 macrophages exposed with SARS-CoV-2 for 48 hours also revealed reduced abundance of proteins primarily associated with PPP, including those related to “oxidoreductase activity” and “NADH metabolic processes”, with another PPP enzyme, 6-Phosphogluconate dehydrogenase, appearing as low- abundant protein. These findings suggest effects of Sars- CoV-2 infection in THP-1 macrophages metabolic pathways. Studies have shown metabolic dysregulation upon Sars- Cov-2 infection in vitro, where SARS-CoV-2 replication in host cells is supported by enhanced aerobic glycolysis and dysregulation in PPP [42, 43]. Regarding the inflammatory response, the analysis highlighted a prominent “inflammatory response to antigenic stimulus,” indicating that the infection elicits an inflammatory effect in THP-1 macrophages, consistent with their innate immune role. The enrichment of “Response to IL-4” and “Response to IL-7” alongside the “IL- 12 mediated signaling pathway” as downregulated pathways indicate modulation of cytokine-mediated signaling. While “response to IL-4” is typically linked to M2-like macrophage polarization (anti-inflammatory/reparative), the “IL- 12 mediated signaling pathway” is associated with Th1 responses and M1-like (pro-inflammatory) polarization.
SARS-CoV-2 employs well described evasion mechanisms, notably suppressing interferon responses [44]. This highlights a dynamic struggle between host recognition and defense systems and the pathogen infection mechanisms. Such an ongoing conflict might explain the absence of an exacerbated pro-inflammatory profile within the proteomic datasets. Additionally, other infected macrophage responses were observed such the downregulation of proteins involved in protein folding, such as Endoplasmin, could further exacerbate endoplasmic reticulum (ER) stress, as evidenced by enriched clusters like “Response to protein unfolding,” “Protein targeting to ER,” and “Protein folding in the ER.” These clusters point towards stress responses related to protein synthesis and folding within the ER, a common consequence of viral infection. SARS-CoV-2 protein synthesis and replication organelle formation induce ER stress. This stress response often involves the overexpression of unfolded protein response (UPR) factors like GRP78, consistently observed at both mRNA and protein levels across various infected cell lines [45, 46, 47] Furthermore, several low-abundant proteins, including Profilin-1, Tropomyosin alpha-4 chain, Gelsolin, F-actin-capping protein alpha-1, Plastin-2, and Vimentin, are crucial components of the cytoskeleton and are essential for cell motility which are vital for an effective immune response and subsequent tissue repair [48].
Conclusion
This study reveals that THP-1-derived macrophages, despite not supporting productive replication of SARS- CoV-2, exposure to the virus and initiates a pronounced inflammatory response. The induction of cytokine secretion and the accompanying proteomic alterations underscore the role of macrophages in shaping the proinflammatory environment associated with COVID-19 pathology. These findings highlight the dual function of macrophages in antiviral defense and in driving immunopathology, offering critical insights into the complex interplay between SARS- CoV-2 and host immune cells and highlight THP-1 cells as a valuable model for investigating viral-host interactions mechanisms.
Conflicts of Interest
The authors declare no conflict of interest.
References
-
WHO COVID19 dashboard. (2025) COVID19 Cases. WHO.
-
Manfrini N, Notarbartolo S, Grifantini R, Pesce E (2024) SARSCoV2: A Glance at the Innate Immune Response Elicited by Infection and Vaccination. Antibodies (Basel) 13(1): 13.
-
Batista JC, DeAntonio R, LópezVergès S (2025) Dynamics of Innate Immunity in SARSCoV2 Infections: Exploring the Impact of Natural Killer Cells, Inflammatory Responses, Viral Evasion Strategies, and Severity. Cells 14(11): 763.
-
Kosyreva A, Dzhalilova D, Lokhonina A, Vishnyakova P, Fatkhudinov T (2021) The Role of Macrophages in the Pathogenesis of SARSCoV2Associated Acute Respiratory Distress Syndrome. Front Immunol 12: 682871.
-
Meidaninikjeh S, Sabouni N, Marzouni HZ, Bengar S, Khalili A, et al. (2021) Monocytes and macrophages in COVID19: Friends and foes. Life Sci 269: 119010.
-
Hoffmann M, Kleine-Weberet H, Schroeder S, Muller MA, Drosten C, et al. (2020) SARSCoV2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 181(2): 271-280.
-
Labzin LI, Chew KY, Eschke K, Wang X, Esposito T, et al. (2023) Macrophage ACE2 is necessary for SARSCoV2 replication and subsequent cytokine responses that restrict continued virion release. Sci Signal 16(782): Eabq1366.
-
Merad M, Martin JC (2020) Pathological inflammation in patients with COVID19: a key role for monocytes and macrophages. Nature Reviews Immunology 20(5): 355- 362.
-
Tay MZ, Poh CM, Rénia L, Ary PAM, Ng LFP (2020) The trinity of COVID19: immunity, inflammation and intervention. Nat Rev Immunol 20(6): 363-374.
-
Schiuma G, Beltrami S, Bortolotti D, Rizzo S, Rizzo R (2022) Innate Immune Response in SARSCoV2 Infection. Microorganisms 10(3): 501.
-
Jiang HW, Zhang HN, Meng QF, Xie J, Li Y, et al. (2020) SARSCoV2 Orf9b suppresses type I interferon responses by targeting TOM70. Cellular & Molecular Immunology 17(7): 998-1000.
-
Ambikan A, Akusjärvi SS, Sperk M, Neogi U (2024) Systemlevel integrative omics analysis to identify the virushost immunometabolic footprint during infection. Adv Immunol 164: 73-100.
-
Pathak AN, Singh LK, Dwivedi E (2021) Omics Approaches for Infectious Diseases. In: Hameed S, Fatima Z (Eds.) Integrated Omics Approaches to Infectious Diseases. Springer, Singapore, pp: 507-519.
-
Li S, Zhang Y, Guan Z, Li H, Ye M, et al. (2020) SARSCoV2 triggers inflammatory responses and cell death through caspase8 activation. Signal Transduct Target Ther 5(1): 235.
-
Codo AC, Davanzo GG, Monteiro De BL, De Souza GF, Muraro SP, et al. (2020) Elevated glucose levels favor SARSCoV2 infection and monocyte response through a HIF1α/glycolysisdependent axis. Cell Metab 32(3): 437- 446.e5.
-
Araujo DB, Machoda RRG, Amgarten DE, Malta FM, De Araujo GG, et al. (2020) SARSCoV2 isolation from the first reported patients in Brazil and establishment of a coordinated task network. Mem Inst Oswaldo Cruz 115: E200342.
-
Hierholzer JC, Killington RA (1996) 2 Virus isolation and quantitation. In: Mahy BWJ, Kangro HO (Eds.) Virology Methods Manual. Academic Press, USA, pp: 25-46.
-
Botosso VF, Jorge SAC, Astray RM, De Sa Guimaraes, Mathor MB, et al. (2022) AntiSARSCoV2 equine F (Ab′)2 immunoglobulin as a possible therapy for COVID19. Scientific Reports 12: 3890.
-
Lund ME, To J, O’Brien BA, Donnelly S (2016) The choice of phorbol 12myristate 13acetate differentiation protocol influences the response of THP1 macrophages to a proinflammatory stimulus. J Immunol Methods 430: 64-70.
-
Manza LL, Stamer SL, Ham AJ, Codreanu SG, Liebler DC (2005) Sample preparation and digestion for proteomic analyses using spin filters. Proteomics 5(7): 1742-1745.
-
Wiśniewski JR, Zougman A, Nagaraj N, Mann M (2009) Universal sample preparation method for proteome analysis. Nat Methods 6(5): 359-362.
-
GiamarellosBourboulis EJ, Netea MG, Rovina N, Akinosoglou K, Antoniadou A, et al. (2020) Complex Immune Dysregulation in COVID19 Patients with Severe Respiratory Failure. Cell Host & Microbe 27(6): 992- 1000.e3.
-
Boumaza A, Gay L, Mezouar S, Bestion E, Diallo AB, et al. (2021) Monocytes and Macrophages, Targets of Severe Acute Respiratory Syndrome Coronavirus 2: The Clue for Coronavirus Disease 2019 Immunoparalysis. J Infect Dis 224(3): 395-406.
-
Atmeh PA, Gay L, Levasseur A, La Scola B, Olive D, et al. (2022) Macrophages and γδ T cells interplay during SARSCoV2 variants infection. Front Immunol 13: 1078741.
-
Zankharia U, Yadav A, Yi Y, Hahn BH, Collman RG (2022) Highly restricted SARSCoV2 receptor expression and resistance to infection by primary human monocytes and monocytederived macrophages. J Leukoc Biol 112(3): 569-576.
-
Junqueira C, Crespo A, Ranjbar S, De Lacerda LB, Lewandrowski M, et al. (2022) FcγRmediated SARSCoV2 infection of monocytes activates inflammation. Nature 606(7914): 576-584.
-
Zhou Y, Fu B, Zheng X, Wang D, Zhoa C, et al. (2020) Pathogenic T cells and inflammatory monocytes incite inflammatory storm in severe COVID19 patients. National Science Review 7(6): 998-1002.
-
Huang X, Zhu W, Zhang H, Qiu S, Shao H (2025) SARSCoV2 N protein induces alveolar epithelial apoptosis via NLRP3 pathway in ARDS. International Immunopharmacology 144: 113503.
-
Grant C, Duffin E, O’Connell F, Nadarajan P, Bergin C, et al. (2025) Human Alveolar Macrophages Detect SARSCoV2 Envelope Protein Through TLR2 and TLR4 and Secrete Cytokines in Response. 175(3): 391-401.
-
Wang Y, Tsai CH, Wang YC, Yen LC, Chang YW, et al. (2024) SARSCoV2 nucleocapsid protein, rather than spike protein, triggers a cytokine storm originating from lung epithelial cells in patients with COVID19. Infection 52(6): 955-983.
-
Chiok K, Hutchison K, Miller LG, Bose S, Miura TA (2023) Proinflammatory Responses in SARSCoV2 and Soluble Spike Glycoprotein S1 Subunit Activated Human Macrophages. Viruses 15(3): 754.
-
Huang H, Li X, Zha D, Lin H, Yang L, et al. (2023) SARSCoV2 E proteininduced THP1 pyroptosis is reversed by Ruscogenin. Biochemistry and Cell Biology 101(4): 303- 312.
-
Pan P, Shen M, Yu Z, Ge W, Chen K, et al. (2021) SARSCoV2 N protein promotes NLRP3 inflammasome activation to induce hyperinflammation. Nature Communications 12(1): 4664.
-
Li W, Liu XJ, Guo YL, Chen CL (2021) SARSCoV2 Nsp5 Activates NFκB Pathway by Upregulating SUMOylation of MAVS. Frontiers in Immunology 12: 750969.
-
Zhang RG, Liu XJ, Guo YL, Chen CL (2024) SARSCoV2 spike protein receptor binding domain promotes IL6 and IL8 release via ATP/P2Y2 and ERK1/2 signaling pathways in human bronchial epithelia. Molecular Immunology 167: 53-61.
-
Patra T, Meyer K, Geerling L, Isbell TS, Hoft DF, et al. (2020) SARSCoV2 spike protein promotes IL6 transsignaling by activation of angiotensin II receptor signaling in epithelial cells. PLOS Pathogens 16(12): e1009128.
-
Baazim H, Koyuncu E, Tuncman G, Burak FM, Merkel L, et al. (2025) FABP4 as a therapeutic host target controlling SARSCoV2 infection. EMBO Molecular Medicine 17(3): 414-440.
-
Cortese M, Lee JY, Cerikan B, Neufeldt CJ, Ooschot VMJ, et al. (2020) Integrative Imaging Reveals SARSCoV2Induced Reshaping of Subcellular Morphologies. Cell Host & Microbe 28(6): 853-866.e5.
-
Roingeard P, Eymieux S, BurlaudGaillard J, Hourioux C, Patient R, et al. (2022) The doublemembrane vesicle (DMV): a virusinduced organelle dedicated to the replication of SARSCoV2 and other positivesense singlestranded RNA viruses. Cellular and Molecular Life Sciences 79(8): 425.
-
Chen ST, Park MD, Valle DMD, Buckup M, Tabachnikova A, et al. (2022) A shift in lung macrophage composition is associated with COVID19 severity and recovery. Science Translational Medicine 14(662): eabn5168.
-
Luzzatto L, Nannelli C, Notaro R (2016) Glucose6Phosphate Dehydrogenase Deficiency. Hematology/Oncology Clinics of North America 30(2): 373-393.
-
Bojkova D, Costa R, Reus P, Bechtel M, Olmer R, et al. (2021) Targeting the Pentose Phosphate Pathway for SARSCoV2 Therapy. Metabolites 11(10): 699.
-
Chen P, Wu M, He Y, Jiang BH, He ML (2023) Metabolic alterations upon SARSCoV2 infection and potential therapeutic targets against coronavirus infection. Signal Transduction and Targeted Therapy 8(1): 237.
-
Kim YM, Shin EC (2021) Type I and III interferon responses in SARSCoV2 infection. Experimental & Molecular Medicine 53(5): 750-760.
-
Basha B, Samuel SM, Triggle CR, Ding H (2012) Endothelial dysfunction in diabetes mellitus: possible involvement of endoplasmic reticulum stress? Experimental Diabetes Research 2012: 481840.
-
Puzyrenko A, Jacobs ER, Sun Y, Felix JC, Sheinin Y, et al. (2021) Pneumocytes are distinguished by highly elevated expression of the ER stress biomarker GRP78, a coreceptor for SARSCoV2, in COVID19 autopsies. Cell Stress & Chaperones 26(5): 859-868.
-
Marano V, Vlachová Š, Tiano SML, Cortese M (2024) A portrait of the infected cell: how SARSCoV2 infection reshapes cellular processes and pathways. npj Viruses 2(1): 66.
-
Wen Z, Zhang Y, Lin Z, Shi K, Jiu Y (2020) Cytoskeleton—a crucial key in host cell for coronavirus infection. Journal of Molecular Cell Biology 12(12): 968-979.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells
- The Role of Faculty in Assessment: Traditional vs. Competency- Based Medical Education in Toxicology Learning