Nectar availability and Interactions with Bird Pollinators in Grevillea banksii R. Br. (Proteaceae) in a Restinga Ecosystem in Southeastern Brazil
The mutualistic relationship between plants and their pollinators may influence species’ spatial distribution, richness, and abundance. We assessed the nectar availability and interaction with pollinators in Grevillea banksii (Proteaceae), an alien plant in Brazil. Our objectives were to study the floral biology and the pollination system and identify the pollinators in a G. banksii population in a Restinga ecosystem (sand-coastal-plain) in southeastern Brazil. We described the behavioral parameters of the pollinator-plant interaction and determined how the pollinators contribute to reproduction. Floral biology was analyzed by taking morphometric measurements of the flowers and determining the distribution of their structures. The reproductive system was assessed via different pollination treatments (open pollination, self-pollination, manual selfpollination, manual cross-pollination). The pollinators were identified and their behaviors were assessed by focal observation. Flower morphometrics showed compatibility between the mean calyx length and the beak length of the flower-visiting hummingbirds. All the pollination types tested were efficient, and we identified eight bird species (Chionomesa fimbriata, Eupetomena macroura, Chlorestes notata, Polytmus guainumbi, Florisuga fusca, Coereba flaveola, Cyanerpes cyaneus and Dacnis cayana) that visit G. banksii in the study area. The results show a positive interaction between pollinators and plants, with offers of resources, pollination, and pollen dispersal.
Introduction
The knowledge of plant floral biology and the pollination system is important to understand the interactions between flowers and their pollinators [1, 2]. This relation between the flower visitor and the plant may be associated with adaptive issues, such as the floral structures having favorable characteristics that facilitate access to flower resources and pollen transport [3].
Plant-pollinator relationship is based on offering a resource to the animal, which, in turn, fertilizes the plant by pollination. In addition to food resources such as nectar and pollen [4], plants also offer volatile oils, resins, shelter, and nest sites to the flower visitors [5, 6, 7]. This mutualistic relation may affect the spatial distribution, richness, and abundance of the different species [8]. Several studies have emphasized the importance of pollination in maintaining species as well as ecosystem characteristics [9]. Therefore, knowledge of the plant-pollinator interaction is important for ecological restoration actions and plant species conservation, as well as for conservation of all the organisms that take part in this process [10].
Some characteristics, in addition to an animal visiting a plant, characterize an animal as the pollinator of a given plant species. The study of visitation frequency and pattern, transport capacity, and quantity of pollen transported may classify an animal as an effective pollinator [11]. Animal preference for native or alien species depends on the quality and quantity of the offered resource. The preference for plant species by nectarivorous birds is determined by the floral attractants retained by each species, such as flower scent, shape, and color [12, 13]. With these characteristics, plant species that are not native to an environment and do not have many predators may reproduce rapidly, thereby attracting more dispersers and suppressing native species [14].
The plant Grevillea banksii belongs to the Proteaceae family and is native from Australia. Its popular names include dwarf silky oak and red silky oak, and it produces a large amount of nectar, attracting many nectarivorous bird species. It has small red flowers placed in pairs and having no petals. Grevillea banksii grows as a shrub and may reach approximately 4 to 6 meters in height [15]. Because of these characteristics, this species was introduced in different countries, including Brazil, as an ornamental plant for landscaping projects [16].
The resources offered by the plants are important for the attraction and maintenance of pollinators, which, in turn, play an essential role in plant reproductive success [13]. Nectar composition and concentration vary among plant species and thus determine the visitation rate and the visitors’ preferences [17, 18]. We thus consider the hypothesis that G. banksii’s reproductive success is affected by the actions of the floral visitors, contributing to the occurrence of cross- pollination in G. banksii, with no self-pollinating individuals in this species.
We assessed the floral biology and pollination system of G. banksii. Our objectives were i) to quantify nectar availability for pollinators; ii) to determine which species pollinate the studied G. banksii population and what type of resource they search for in the flowers; iii) to determine the behavioral parameters and the pollinator-plant interactions and; iv) to determine how the pollinators contribute to the plants’ reproduction.
Material and Methods
Study Site
The study was conducted in the Setiba Environmental Protection Area (Área de Proteção Ambiental de Setiba, hereafter APA-Setiba), which includes the Paulo César Vinha State Park (Figure 1). The APA-Setiba is located in the Guarapari municipality, state of Espírito Santo, southeastern Brazil, and encompasses 5,460 hectares of marine area and 7,500 ha of terrestrial land between the coordinates 20°33’ – 20°38’ S and 40°23’ – 40°26’ W [19]. According to the Köppen classification, the regional climate is monsoon (Am) [20] with hot and rainy summers and dry winters, a mean annual temperature of 23.3°C, a mean annual rainfall of 1,307 mm, and a mean annual relative humidity of 80% [21].
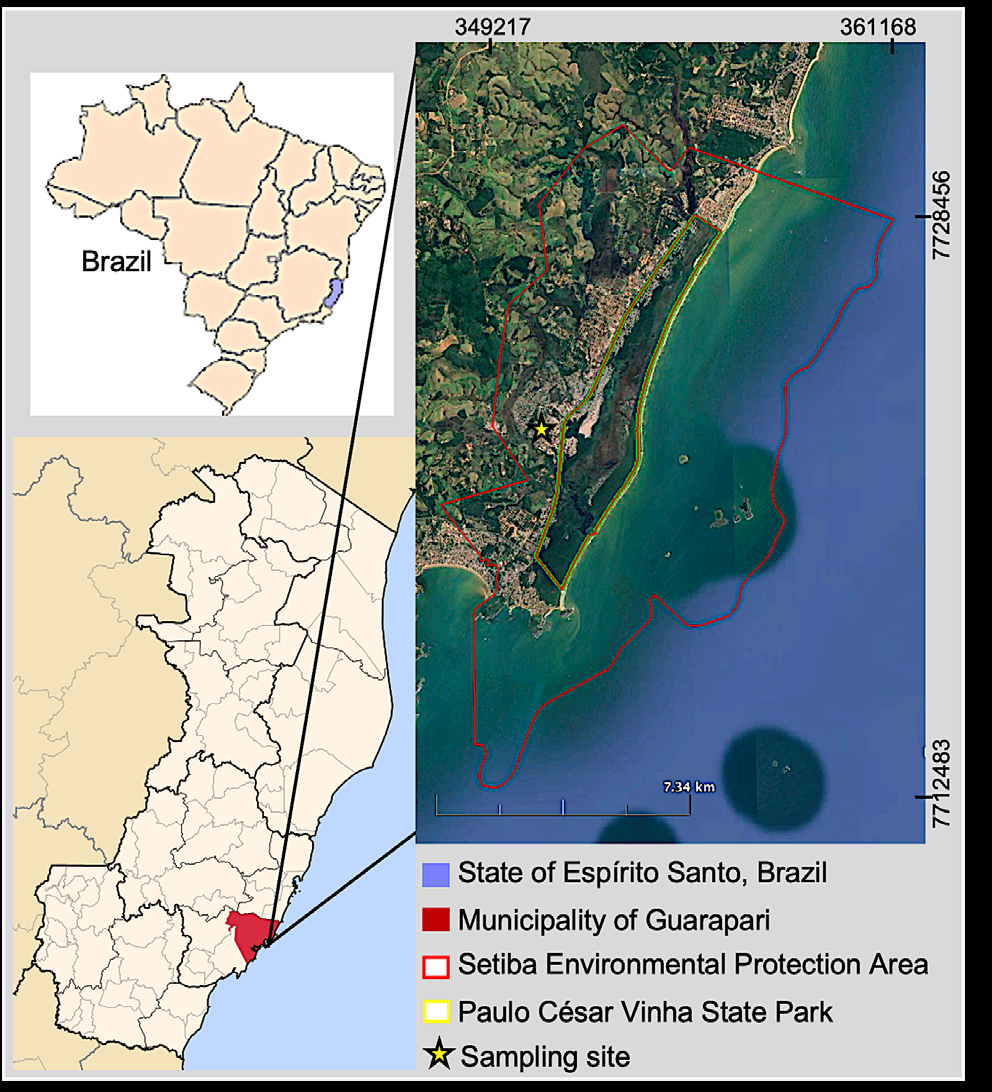
Figure 1: Location of sampling site in Setiba Environmental Protection Area, relative to municipality of Guarapari in state of Espírito Santo, southeastern Brazil. The APA-Setiba is located in the Atlantic Forest biome, specifically in the esting ecosystem (sand-coastal-plain). The APA-Setiba is crossed by the highway ES-060 (Rodovia do Sol) and is surrounded by residences, farms, and pastures. A large number of people access the area, contributing to its habitat degradation. In the past, sand extraction was one of the activities that most contributed to environmental degradation, resulting in fragments with landscape changes. The studied G. banksii population is located mostly in the degraded portions of the APA-Setiba at 3.3 m asl. The field sampling was done between December 2014 and August 2015, starting in the spring and finishing in the winter.
Flower Biology and Morphometry
The floral biology aspects of G. banksii were studied by determining the flower structures and their distribution. We collected 30 flowers from different individuals and used a digital caliper to measure the calyx diameter and length, stigma and style diameter, and length from stigma to androecium (Figure 2). The flower structures were then placed on millimeter paper and photographed. These observations provided data on flower morphology, which are important to understand the interaction between the animal and the flower parts.
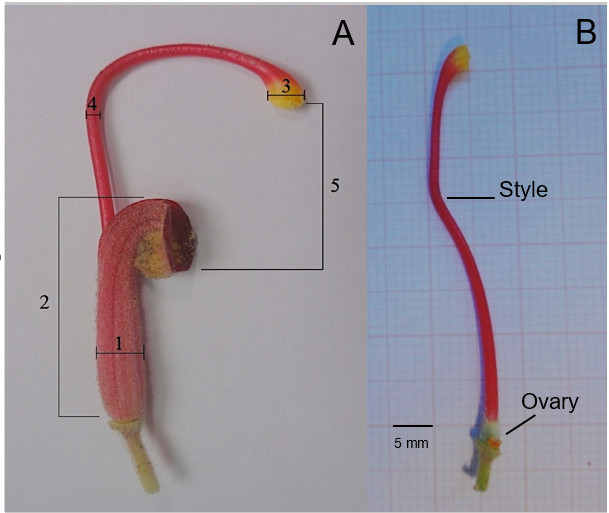
The average calyx length of G. banksii flowers was compared to the average beak length of the flower-visiting birds (see flower visitors). To estimate the average number of flowers on G. banksii inflorescences, 30 shrubs were selected, and the flowers of one inflorescence were counted on each of these shrubs.
Reproductive System
To determine and understand the pollination system of G. banksii, 12 plants were tagged, and the following controlled pollination treatments were applied: i) open pollination, where the marked inflorescence remained open and susceptible to pollinator visits; ii) autonomous self-pollination, where the inflorescences were bagged with voile and remained bagged until fruiting; iii) manual self-pollination, where the bagged inflorescences were manually pollinated with pollen from other flowers from the same individual; and iv) manual cross-pollination, where the bagged inflorescences were manually pollinated with pollen from other individuals. After fruiting, the bagged inflorescences were opened to count the fruits formed. This enabled a determination of the plant’s reproductive success according to the number of seeds produced and the acquisition of information on the plant’s pollination system.
Nectar Flow
Nectar flow was assessed in an experiment with five nearby individuals. Two inflorescences from each of these individuals were subjected to two different treatments: open and closed systems. In the closed system, the inflorescences were bagged with voile, precluding the access to the flowers. In the open system, the inflorescences were only tagged for posterior identification and pollinator access to the flowers was not restricted. We thus had five inflorescences for each treatment (closed and open system). Nectar was collected from the flowers three times per day (at 06h30min am, 11h30min am, and 4h30min pm) with microcapillary tubes that were introduced to the flower’s calyx. In each collecting period, one flower from each of the five inflorescences in the two treatments was used for nectar collection, as the flowers fell from the inflorescence during this process. The space occupied by the nectar in the microcapillary tube was measured with a digital caliper for volume calculation. Solute concentration was measured with a pocket refractometer (Model BTX-1, Vee Gee Brix, range of 0-32%). Nectar volume was calculated by multiplying the total volume of the microcapillary tube (70 μL) by the length occupied by the nectar (mm) in the tube and dividing it by the total length of the microcapillary tube (75 mm). During nectar collection, the air temperature and moisture were also recorded with a digital thermo-hygrometer (Minipa MT-240). The data obtained in this experiment showed the differences in nectar volume between the closed and open systems, and the variation in the quantity of this resource at different periods of the day.
Flower Visitors
The flower-visiting birds and their behavior during the visits were recorded via mist net captures and focal individual observation. For focal individual observation, 10 areas were delimited; and three to six plants were observed simultaneously in each of these areas. These observations amounted to 50 observation hours taken over the course of 5 days and performed from 6h30min am to 4h30min pm each day. The following information was recorded during the observations: i) bird species and behavior during the visit; ii) the resource searched for by the animal in the flower (nectar and/or pollen); iii) number of visited inflorescences and flowers and; iv) observation time and duration.
For sampling of the flower visitors by capture, 10 mist nets were placed in two different areas close to G. banksii individuals to capture the birds that would probably be visiting the plants. The birds were identified based on the literature, and the presence of any sort of pollen on the bodies of the captured individuals was also noted. The beak length of each captured individual was obtained according to Sick [22].
Data Analysis
One-way ANOVA was used to assess the possible difference in reproductive success among the pollination treatments and nectar volume and nectar concentration of inflorescences of the closed and open systems. Spearman correlation was used to assess whether there was a correlation between nectar volume and concentration for inflorescences of the closed and open system inflorescences. The same analysis was used to assess the relation between nectar volume and temperature and moisture. A one-way ANOVA followed by the Tukey post-hoc test was used to assess whether nectar volume and concentration varied throughout the day. All these analyses were performed in the BioEstat 5.0 software [23], with a 5% significance level.
Results
Flower Biology
Grevillea banksii flowers are stacked and presented in pairs forming terminal inflorescence clusters (Figure 3). The flowers are of an intense red color, and the anthers and stigmas are yellowish. The nectaries are located within the calyx, which serves as a reservoir when the nectar is secreted. The calyx also contains the flower’s ovary and, attached to it, the style, which is a long extension that branches towards the outside of the calyx (Figure 2). This forms a concave projection, originating a chamber that contains the anthers (Figure 2).

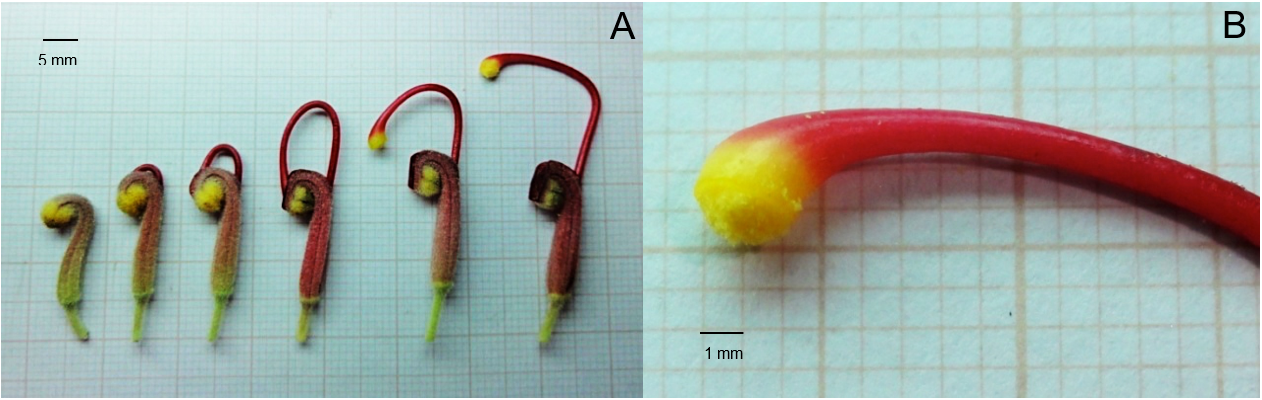
The style remains curved, with the stigma being located close to the chamber with the anthers (Figure 4A). When shedding, the stigma becomes filled with pollen grains and functions as a deposit (Figure 4B). The style loses its curve and becomes more distant from the anther, resulting in an angle between the stigma and the calyx’s cavity (Figure 4A) and permitting the flower visitor to access the nectar. When collecting this resource, a part of the animal’s body touches the pollen-impregnated stigma and transports it to other flowers.
Grevillea banksii fruits are grayish-green and hairy (Figure 5A). Mature fruits are brown and dry by opening in the middle (Figure 5B). Up to two seeds are formed within the fruit cavity. The seeds are flat and covered by a coat that is less thin at the extremities (Figure 5C).

The results of the morphometric data for G. banksii inflorescences (Table 1) show the compatibility between the average calyx length and the beak of some hummingbird species that visited the flowers (Table 2).
| Floral Attributes (mm) | Average ± SD |
| Calyx diameter | 3.32 ± 0.24 |
| Calyx length | 18.37 ± 0.70 |
| Stigma diameter | 1.95 ± 0.14 |
| Style diameter | 0.73 ± 0.07 |
| Length from stigma to androecium | 14.8 ± 3.51 |
Table 2: Measurements of the floral attributes of Grevillea banksii (mm) with their respective averages and standard deviations (
| Species (n) | Beak Length (mm) |
|---|---|
| Chionomesa fimbriata (n = 30) | 19.3 ± 2.1 |
| Eupetomena macroura (n = 30) | 19.6 ± 1.3 |
| Polytmus guainumbi (n = 30) | 21.2 ± 1.4 |
| Chlorestes notata (n = 19) | 16.0 ± 1.6 |
| Florisuga fusca (n = 1) | 19.6 |
| Coereba flaveola (n = 30) | 9.5 ± 1.6 |
| Dacnis cayana (n = 16) | 9.2 ± 0.7 |
| Cyanerpes cyaneus (n = 20) | 13.0 ± 1.0 |
Table 1: Bird species recorded as flower visitors of Grevillea banksii and average beak length (mm) ± standard deviation of each
The beak lengths of the hummingbirds Chionomesa fimbriata, Eupetomena macroura, Polytmus guainumbi, Chlorestes notata, and Florisuga fusca (Table 2) are compatible with the average calyx length of the G. banksii flowers (18.37 ± 0.70).
Pollination System
The results obtained in the pollination-system experiment show that all pollination types tested were efficient. Even though the fruit quantity varied, the number of seeds per fruit was the same among all treatments (Table 3). The average number of flowers per G. banksii inflorescence was 39 ± 8.6.
| Treatment | Flowers (n) | Fruits (n) | Seeds (n) | Reproductive Success (%) |
|---|---|---|---|---|
| Open pollination | 39 | 0 | 0 | 26.90% |
| 39 | 16 | 32 | ||
| 39 | 5 | 10 | ||
| Self-pollination | 39 | 15 | 30 | 24.80% |
| 39 | 8 | 16 | ||
| 39 | 6 | 12 | ||
| Manual self-pollination | 39 | 16 | 32 | 33.30% |
| 39 | 5 | 10 | ||
| 39 | 18 | 36 | ||
| Cross-pollination | 39 | 14 | 28 | 17.10% |
| 39 | 1 | 2 | ||
| 39 | 5 | 10 |
Table 3: Results of the pollination treatment and average reproductive success (%) of Grevillea banksii at the Setiba Environment
Reproductive success was determined as the difference between the number of fruits and of flowers, with no significant differences being observed for the reproductive success values between the pollination treatments (F = 0.566; d.f. = 3; P = 0.655).
Nectar Flow
Total nectar volumes during the sampling day were significantly different between the closed and open systems (F = 13.632; d.f. = 1; P = 0.001). During the collection hours, the inflorescences of the closed system had a greater total volume (26.96 µL) than those of the open system (13.94 µL) (Table 4). No significant differences were observed in the nectar solute concentration between the inflorescences of the closed and open systems (F = 0.064; d.f. = 1; P = 0.797) (Table 4).
| Hour | Temperature (°C) / Humidity | Open System | Closed System | ||
|---|---|---|---|---|---|
| Volume (µL) | Concentration (% Brix) | Volume (µL) | Concentration (% Brix) | ||
| 06h30min | 21.1°C / 74% | 20.95 ± 14.04 | 14.04 ± 2.40 | 24.62 ± 14.51 | 17.40 ± 4.75 |
| 11h30min | 33.7°C / 36% | 9.58 ± 4.23 | 18.14 ± 3.85 | 27.83 ± 8.24 | 16.24 ± 4.20 |
| 16h30min | 25.2°C / 53% | 11.29 ± 5.47 | 20.84 ± 3.60 | 28.43 ± 5.00 | 18.28 ± 2.13 |
Table 4: Average and standard deviation of nectar volume and concentration obtained for Grevillea banksii inflorescences in close
Nectar volume and concentration were not positively correlated in either the open-system (rs = -0.268; d.f. = 14; P = 0.332) or the closed-system inflorescences (rs = -0.055; d.f. = 14; P = 0.844). No significant correlations were observed between the nectar volume and the moisture (rs = -0.170; d.f.
= 14; P = 0.544) and temperature (rs = 0.170; d.f. = 14; P = 0.544) for the closed-system inflorescences. Similarly, no significant correlations were observed between the nectar volume and moisture (rs = 0.492; d.f. = 14; P = 0.062) and temperature (rs = -0.492; d.f. = 14; P = 0.062) for the open-system inflorescences.
No significant variation was observed in the nectar volume throughout the day for either the open-system (F = 2.297; d.f. =2; P = 0.141) or the closed-system inflorescences (F = 0.207; d.f. = 2; P = 0.816).
No significant variation was observed in the nectar concentration throughout the day for the closed-system inflorescences (F = 0.351; d.f. = 2; P = 0.714). However, in the open-system inflorescences, nectar concentration varied between the sampling hours (F = 5.243; d.f. = 2; P = 0.022), with significant differences only between the first and the last nectar sampling times (Tukey post-hoc P < 0.05).
Flower Visitors
G. banksii flowers were visited by birds and insects (mostly bees) that searched for nectar as a resource. In each visit, the birds visited three to five inflorescences per shrub and an average of three flowers per inflorescence. The birds hovered in the air and inserted their beaks into the flower’s calyx. This process lasted between 2 and 6 seconds, after which the bird visited another flower on the same inflorescence. The birds usually visited more than one nearby shrub per flight, landed for several seconds, and then either moved to another area or continued the visitations in the same area.
A total of 84 individuals belonging to eight bird species were captured as they visited the G. banksii shrubs. The species C. fimbriata (n = 34) was the most captured species in the area, followed by E. macroura (n = 16), (Figures 6A), Coereba flaveola (n = 12), Cyanerpes cyaneus (n = 8), P. guainumbi (n = 6), C. notata (n = 4), (Figure 6B), Dacnis cayana (n = 3), and F. fusca (n = 1). Fifty seven percent (n = 48) of the captured birds showed no signs of pollen deposition on any part of their bodies. However, 16 individuals of C. fimbriata and, eight of E. macroura and three of C. flaveola had pollen vestiges on the upper maxilla, and signs of pollen depositions were observed on the crown of nine individuals belonging to different species (C. fimbriata, C. flaveola, F. fusca, and E. macroura).

Discussion
Flower Biology and Morphometry
The set of flower measurements (calyx length and length from stigma to androecium) provides important information on plant-pollinator interactions. Importantly, the G. banksii flower characteristics may contribute to the success of pollinator visits for pollination and pollen dispersal. According to the obtained measurements, the beak lengths of the species C. fimbriata, P. guainumbi, C. notata, and E. macroura are equivalent to the G. banksii calyx length. This morphological similarity favors the pollinator that inserts its beak into the flower’s calyx to collect the nectar and benefits the plant by facilitating pollen deposition on the animal’s body and enabling its dispersal. Pollen dispersal and pollination are favored by the style’s curvature and the location of the pollen-impregnated stigma.
Pollinator visits and the variety of pollinators visiting a flower are defined by the pollinators’ characteristics. Plants may vary in their calyx lengths, number of flowers per inflorescence, population density, and nectar quantity [24]. G. banksii has many flowers per individuals, and the individuals are numerous and found in patches. This clustered pattern may be related to a larger number of visitors, as the number of visits and the variety of visitors are related to the establishment of a plant species in the area and to its survival rate. These characteristics are also related to the flowers’ adaptation to different types of pollinators. This is especially true for alien species, which may not have effective pollinators in their establishment area [25].
Pollination System
Some plants have different distances and directions between the anther and the stigma, precluding self- pollination [26]. Notwithstanding the physical difference between the mature anther and stigma, the stigma had been in physical contact with the androecium, enabling pollen deposition on the style’s extremity where the stigma is located. This pollen deposition may be related to the positive result of the self-pollination treatment.
The lack of significant differences between the pollination treatments showed that G. banksii is benefited by both cross- pollination and self-pollination. Similar results were obtained for Momordica charantia (Cucurbitaceae) [27] and Cucurbita moschata flowers (Cucurbitaceae) [28] and Psittacanthus robustus (Loranthaceae) [13]. Even though self-pollination is efficient in G. banksii, a large amount of resources is offered to the visitors, and the flower characteristics are attractive to the pollinators. These characteristics favor cross-pollination by attracting pollinators and thus ensure the genetic variability in the population [24].
No significant differences were observed in the species’ reproductive success among the pollination treatments, showing that all pollination types are efficient. This result shows that G. banksii pollinators in the study area have the same contribution to offspring production as self-pollination. This may be explained by a lack of coevolution between pollinators and G. banksii in the study area, as G. banksii is not native to Brazil, and all the flower visitors belong to the Brazilian bird fauna. Notwithstanding, pollinator activity is important for the species because it ensures the population’s genetic variability.
Nectar Flow
The nectar concentration in G. banksii did not vary throughout the day for the closed inflorescences. A constant nectar concentration has also been observed for Geoffroea decorticans (Fabaceae) [29] during daytime. On the other hand, the inflorescences that remained susceptible to visitors showed a difference in nectar concentration between the first and last hour of pollinator visits (6h30min am: 14,4%, 16h30min pm 20.8%). Daily variation in the nectar concentration was also observed for Plumbago auriculata (Plumbaginaceae) with smaller concentrations in the morning [30], and for P. robustus the amount of nectar per flower was highest at beginning of anthesis [13]. Sugar concentration can also be decreased throughout the day by flower mites action, reducing the amount of sugar available to pollinators [31].
Variation in nectar concentration during the day may be related to hummingbird visits to G. banksii inflorescences, as visitor activity peaked mostly between 6h30min am and 11h00min am. Nectar concentration was thus smaller during the first hours of the day and increased after the birds collected this resource from the flowers. Some ornithophilous species present nectar replenishment after visit of nectarivorous birds [18].
The lower nectar volume in the open-system inflorescences was probably due to the birds’ and insects’ visiting the flowers and collecting the available resource. Differences in nectar volume between closed-system and open-system flowers have also been observed in Palicourea crocea [32] and Rubus sp. [33]. These authors also ascribed the discrepancy in nectar volume to the high visitation rate of nectar-seeking pollinators. The smaller nectar volume in open-system flowers was also associated with the visits of pollinators searching for resources, especially in the morning [27]. These results may indicate strong interactions between the visitors’ behavior and the patterns of nectar secretion by the visited plants, showing that, even though the study species is an alien one, it established a tight mutualistic relation with the native pollinator species.
Flower Visitors
The birds made fast visits to the inflorescences and then moved to farther patches or remained in the same patch continuing to forage on nearby individuals, but always switching among inflorescences and shrubs. This type of brief visit alternates among plants, favoring cross-pollination [34, 35], and is thus considered to be the most efficient for the plants when compared to visitation by animals that spend more time on the same shrub [36]. It is also favorable to the animal that needs nectar to supply its energy needs and thus visits numerous inflorescences that provide the sought-after resource.
Visits to G. banksii inflorescences occurred during the entire observation time, but higher bird activity was observed between 6h30min am and 11h00min am. A larger volume of nectar produced by the plants was also recorded during this period. Thus, a relation between visitation and nectar production is possible, with pollinators being able to adapt their foraging behavior to the time period with the greatest nectar availability. The synchrony between the time with the greater visit frequency and the period with the greatest volume of produced nectar has also been observed for other species [37, 38].
All recorded bird species may be pollinators of G. banksii, as all were observed visiting the flowers of this species and were captured next to G. banksii individuals. Evidence also existed of pollen accumulation on bird bodies and of nectarivorous feeding habits in these species.
Coereba flaveola is known by its habit of piercing the flowers’ calyx in search for nectar, and it acts as a pollinator of bird-pollinated species [39, 40]. Chionomesa fimbriata has also been described as nectarivorous and with morning foraging habits [24], and E. macroura was considered as main pollinators of P. robustus [13]. Chionomesa fimbriata, C. notata, F. fusca, E. macroura, and P. guainumbi belong to the Trochilidae family and have nectarivorous feeding as a basic characteristic [22]. Cyanerpes cyaneus and D. cayana are Passeriformes that eat plants (fruits, leaves, buds, and nectar) as part of their diet [22, 41].
Little variation was observed in the position of the mature stigma among the G. banksii flowers, and consequently, pollen accumulation was observed on only two areas of the birds’ bodies (crown and dorsal maxilla). Pollen is therefore markedly deposited on the same body parts, favoring the transport of large quantities of pollen. In addition, this occurrence of pollen on these parts of the body may also increase the probability of the transported pollen contacting the stigma of the other flowers when the bird visits other inflorescences. The combination of these processes results in greater pollen deposition on these areas, suggesting that pollination is efficient and that the pollinators contribute to plant reproduction. In addition, plant deposition on the crown or the maxilla appears to be more advantageous, as these areas hamper the loss of pollen grains during flight [42].
Acknowledgments
CD is grateful to CNPq (Bolsa de Produtividade em Pesquisa, grant 308997/2023-9). We thank IEMA (Instituto Estadual do Meio Ambiente) for sampling permit. M. L. Garbin and L. D. Thomaz for suggestions that improved the manuscript and T. J. A. Guerra for helping in methods and data collecting. B. H. Paixão received scholarships from FAPES (Foundation for the Support of Research of Espírito Santo).
Running Title
Interactions with bird pollinators in Grevillea banksii
References
-
Galetto L, Bernardello L (2004) Floral Nectaries, Nectar Production Dynamics and Chemical Composition in Six Ipomea Species (Convolvulaceae) in Relation to Pollinators. Ann Bot 94(2): 269-280.
-
Agostini K, Sazima M, Galetto L (2011) Nectar Production Dynamics and Sugar Composition in Two Mucuna Species (Leguminosae, Faboideae) with Different Specialized Pollinators. Naturwissenschaften 98(11): 933-942.
-
Zen DM, Acra LA (2005) Biologia Floral E Reprodutiva De Agapanthus Africanus (L.) Hoffmanns (Liliaceae). Estudos Biol 27(59): 35-39.
-
Cushman JH, Beattie A (1991) Mutualisms: Assessing the Benefits to Hosts and Visitors. Trends Ecol Evol 6(6): 193-195.
-
Simpson BB, Neff JL (1983) Evolution and Diversity of Floral Rewards. In: Jones CE, Little RJ (Eds.), Handbook of Experimental Pollination Biology. Scientific and Academic Editions, New York, pp: 142-159.
-
Gottsberger G (1984) Pollination strategies in Brazilian Philodendron Species. Ber Deut Bot Ges 97: 391-410.
-
Prance GT (1985) The Pollination of Amazonian Plants. In: Prance GT, Lovejoy TE (Eds), Key environments: Amazonia. Oxford Press, New York, pp: 166-191.
-
Heithaus ER (1974) The Role of Plant-Pollinator Interactions in Determining Community Structure. Ann Mo Bot Gard 61: 675-691.
-
Potts SG, Biesmeijer JC, Kremen C, Neumann P, Schweiger O, et al. (2010) Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol Evol 25(6): 345-353.
-
Bawa KS (1990) Plant-pollinator interactions in tropical rain forests. Annu Rev Ecol Syst 21: 399-422.
-
Stone JL (1996) Components of Pollination Effectiveness in Psychotria Suerrensis, a Tropical Distylous Shrub. Oecologia 107: 504-512.
-
Toledo MCB, Donatelli RJ (2010) Spectral Analyses of Flowers Used by Nectar-Feeding Birds in an Urban Area in Southeastern Brazil. Braz J Biol 70(S3): 729-735.
-
Guerra TJ, Galetto L, Silva WR (2014) Nectar Secretion Dynamic Links Pollinator Behavior to Consequences for Plant Reproductive Success in the Ornithophilous Mistletoe Psittacanthus Robustus. Plant Biol 16(5): 956- 966.
-
French K, Major R, Hely K (2004) Use of Native and Exotic Garden Plants by Suburban Nectarivorous Birds. Biological Conservation 121(4): 545-559.
-
Silva DB, Lemos BS (2002) Plants in the green area of Super Quadra 416 Norte - Brasilia, DF. EMBRAPA 1: 1-147.
-
Pinheiro MS, Sato KA, Dias LP, Renó FGF, Souza TGG, et al. (2010) Effects of pollution on the phenology of Grevillea banksii in the municipality of São José dos Campos-SP. In XIV Latin American Meeting of Scientific Initiation and X Latin American Meeting of Postgraduate Studies. University of Vale do Paraíba, São Paulo, pp: 1-4.
-
Krömer T, Kessler M, Lohaus G, Schmidt-lebuhn AN (2008) Nectar sugar Composition and Concentration in Relation to Pollination Syndromes in Bromeliaceae. Plant Biol 10(4): 502-511.
-
Stahl JM, Nepi M, Galetto L, Guimarães E, Machado SR (2012) Functional Aspects of Floral Nectar Secretion of Ananas Ananassoides, an Ornithophilous Bromeliad from the Brazilian Savanna. Ann Bot 109(7): 1243-1252.
-
Martins MLL, Carvalho-Okano RM, Luceño M (1999) Cyperaceae do Parque Estadual Paulo César Vinha, Guarapari, Espírito Santo, Brasil. Acta Bot Bras 13(2): 187-222.
-
Alvares CA, Stape JL, Sentelhas PC, Gonçalves JLM, Sparovek G (2013) Köppen’s climate classification map for Brazil. Meteorologische Zeitschrift 22(6): 711-728.
-
Fabris LC (1995) Floristic and phytosociological composition of a strip of coastal sandy forest in the Setiba State Park, Municipality of Guarapari, ES. MSc. Dissertation, São Paulo: Universidade Estadual Paulista.
-
Sick H (2001) Brazilian Ornithology. Editora Nova Fronteira 3rd (Edn.), Rio de Janeiro.
-
Ayres M, Ayres-Jr M (2000) BioStat 3.0: Statistical Applications in the Areas of Biological and Medical Sciences. Civil Society of Mamiraua, Belem, CNPq, Brasilia.
-
Locatelli E, Machado IC, Medeiros P (2004) Saranthe klotzschiana (Koer.) Eichl. (Marantaceae) E Seu Mecanismo Explosivo De Polinização. Rev Brasil Bot 27(4): 757-765.
-
Baker HG (1961) The Adaptation of Flowering Plants to Nocturnal and Crepuscular Pollinators. Q. Rev Biol 36(1): 64-73.
-
SanMartin-Gajardo I, Sazima M (2005) Espécies De Vanhouttea Lem. E Sinningia Nees (Gesneriaceae) Polinizadas Por Beija-Flores: Interações Relacionadas Ao Hábitat Da Planta E Ao Néctar. Rev Bras Bot 28(3): 441-450.
-
Lenzi M, Orth AI, Guerra TM (2005) Ecologia Da Polinização De Momordica Charantia L. (Cucurbitaceae), em Florianópolis, SC, Brasil. Rev Brasil Bot 28(3): 505- 513.
-
Cardoso AII (2003) Produção E Qualidade De Sementes De Abobrinha “Piramoita” Em Resposta À Quantidade De Pólen. Bragantia 62(1): 47-52.
-
Eynard C, Galetto L (2002) Pollination Ecology of Geoffroeae Decorticans (Fabaceae) in Central Argentine Dry Forest. J Arid Environments 51(1): 79-88.
-
Galetto L (1993) El Nectário y la Composicion Quimica del Nectar en Plumbago auriculata y P. caeruela (Plumbaginaceae). B Soc Argent Bot 29: 67-72.
-
Guerra TJ, Romero GQ, Benson WW (2010) Flower Mites Decrease Nectar Availability in the Rain-Forest Bromeliad Neoregelia Johannis. J Trop Ecol 26(4): 373- 379.
-
Mendonça LB, Anjos L (2006) Flower Morphology, Nectar Features, and Hummingbird Visitation to Palicourea Crocea (Rubiaceae) In the Upper Paraná River Floodplain, Brazil. An Acad Bras Cienc 78(1): 45-57.
-
Junior, LJM (2007) Ecologia da Polinização de Amoreira- Preta (Rubus sp.) (Rosaceae) em Timbó, SC, Sul do Brasil. MSc. Dissertation, Florianópolis: Universidade Federal de Santa Catarina.
-
Canela MBF, Sazima M (2003) Florivory by the Crab Armases Angustipes (Grapsidae) Influences Hummingbird Visits to Aechmea Pectinata (Bromeliaceae). Biotropica 35(2): 289-294.
-
Perez-Navarro ML, Lopez J, Rodriguez-Riano T, Bacchetta G, Gordillo CM, et al. (2017) Confirmed Mixed- Insect Pollination System of Scrophularia Trifolia L., a Tyrrhenian Species with Calyx Spots. Plant Biol 19(3): 460-468.
-
Gill FB (1985) Hummingbird flight speeds. Auk 102: 97- 101.
-
Fischer E, Leal IR (2006) Effect of Nectar Secretion Rate on Pollination Success of Passiflora Coccinea (Passifloraceae) in the Central Amazon. Braz J Biol 66(2B): 747-754.
-
Longo ML, Fischer E (2006) Efeito da taxa de secreção de néctar sobre a polinização e a produção de sementes em flores de Passiflora speciosa Gardn. (Passifloraceae) no Pantanal. Rev Brasil Bot 29(3): 481-488.
-
Neill DA (1987) Trapliners in the Trees: Hummingbird Pollination of Erythrina Sect. Erythrina (Leguminosae: Papilionoideae). Ann Missouri Bot Gard 74: 27-41.
-
Snow BK, Snow DW (1971) The Feeding Ecology of Tanagers and Honeycreepers in Trinidad. Auk 88: 291-
-
Lopes LE, Fernandes AM, Marini MÂ (2005) Diet of Some Atlantic Forest Birds. Rev Bras Ornitol 13(22): 95-103.
-
Silva CA, Vieira MF, Amaral CH (2010) Floral Attributes, Ornithophily and Reproductive Success of Palicourea Longepedunculata (Rubiaceae), a Distylous Shrub in Southeastern Brazil. Rev Bras Bot 33(2): 207-213.
- Lessons to Learn: Trees are More than the Lungs of the World
- Community Forestry Enterprises as a Model for Sustainable Forest Development: The Case Of The "Baja Tarahumara" in Chihuahua, Mexico
- Ecological and Socio-Economic Impacts of Chromolaena odorata and Mesosphaerum suaveolens, Two Invasive Alien Species in Central and Southern Benin, West Africa
- Epigenetic Sustainability: Modeling the Human Factor as a Natural Resource through Science 4.0 and the NR3C1 Biological Pilot
- Growth-at-Risk: A Framework for Assessing Economic Vulnerability
- The Rural Territory as a Socioecological System for the Management of Public Policy for Sustainable Rural Development