Evaluation of the Allelopathic Activity of Mimosa pudica (L.) Weed Extracts on the Germination of Okra Seeds
The objective of this study was to evaluate the allelopathic effect of Mimosa pudica (L.) on okra germination. The experiment was carried out in the laboratory, where the seeds (20/ Petri dish) of two okra varieties (Clemson: V1 and Hire: V2) were watered with different concentrations (C1: 25%, C2: 50%, C3: 75%, C4: 100%) of aqueous extracts from the roots (RAE), stems (SAE), and leaves (LAE) of Mimosa. Control seeds were watered with sterile distilled water (C0: 0%). Germination parameters such as average growth rate, coefficient of velocity, duration of germination, germination energy, germination%, germination vigor index, germination index, inhibition, last day of germination, latency time, radicle length, mean daily germination, mean germination time, phytotoxicity, seed germination index, speed of emergence, speed of germination, and time to 50% germination were evaluated. The aqueous extracts of Mimosa negatively affected the germination parameters, with effects being concentration-dependent. At V2, the highest germination percentage was recorded for the control (C0: 81.67%), while the lowest was observed at C4: 6.67% for LAE. In V1, radicle length was 5.77, 0.62, 0.18, 0, and 0 cm for C0, C1, C2, C3, and C4, respectively, for SAE. The seeds of V2 were less affected by aqueous extracts compared to V1, with inhibition recorded at 60.64% in V1C1 versus only 19.94% in V2C1. Inhibition was significantly stronger with LAE and SAE compared to RAE. From C1 to C4, AE phytotoxicity ranged from 0.005 to 0.01 for LAE, from 0.005 to 0.01 for SAE, and from 0.001 to 0.007 for RAE. The burial of this weed is therefore prohibited. These findings show that Mimosa aqueous extracts contain toxic ingredients and suggest that LAE and SAE should be tested on weeds to assess their bioactivity, as they can serve as a basis for bioherbicides.
Introduction
Okra (Abelmoschus esculentus (L.) Moench) is a unique vegetable crop, as every part of the plant is used in medicine, human nutrition, industry (glossy paper, sizing agent), and crafts [1, 2, 3]. It contains high levels of calcium, iron, protein, vitamins (A, B6, B9, and C), phosphorus, magnesium, and fiber, nutrients that can help address deficiencies and reduce malnutrition in poor countries worldwide [4]. The crop is highly profitable for growers. However, Cameroon’s annual production remains low (77,631.82 tons) compared to that of other countries in the sub-region, such as Nigeria (1,911,818.50 tons) [5].
Cameroon’s production increased steadily from 1990 (10,000 tons) to 2016 (78,763 tons) [5]. However, since 2017 (69,060 tons), a significant decline in production has been observed at both national and regional levels, and production has not returned to its 2016 peak. Biotic and abiotic constraints are generally responsible for this decline in production [6, 7]. Among the main biotic constraints is allelopathy which is the chemical inhibition of one plant by another. The interaction between grass growth and plant growth leads to yield losses due to weeds. These losses are caused by allelochemicals, which are toxic to plant germination and growth [8].
Mimosa pudica L. is an invasive species that inhibits the germination and growth of other plants, such as okra. High levels of allelochemicals have been detected in its leaves, pods, and seeds [9]. Although M. pudica is a widespread invasive plant in Cameroon, to our knowledge, no studies have been conducted on its allelopathic effect on seed germination and crop growth. Hence, this study aims to assess the allelopathic effect of aqueous extracts from M. pudica leaves, stems, and roots on okra seed germination.
Material and Methods
Study Sites
Weeds were collected from the experimental farm of the Faculty of Sciences (FS), University of Yaoundé I (UYI) (Geographic coordinates: 3°51′30″N-11°29′30″E, altitude: 710 m). The average annual temperature of the site is 24°C, with precipitation ranging from 1,000 to 2,000 mm/year. There is a four-season climate, with two rainy seasons (March to June: short season, September to November: long season) and two dry seasons (July to August: short season, December to February: long season).
The soil is sandy-loam with 34.5% sand, 35% silt, and 30.5% clay [10]. The phytotoxicity test was carried out at the Plant Biotechnology Laboratory of FS-UYI.
Plant Materials
The plant material consisted of seeds from two varieties of okra. The first was Clemson with smooth and angular fruits. It is vigorous, early (50-55 days), and heat-demanding. The second was the Hire variety, a hybrid adapted to high temperatures, early (60-65 days). Both varieties were purchased at the Mfoundi seed market (Centre Region).
The weed consisted of the organs (leaves, stem, and root) of Mimosa pudica.
Methods
Fresh green leaves, fresh green stems, and mature roots of M. pudica were first harvested. Each organ was washed with tap water and rinsed with distilled water. Then, the plant materials were dried at 48 °C in an oven for 72 hours and ground to obtain a powder with a fine and uniform texture. The powders of each sample were stored in a polyethylene bag until use.
Leaf (LEA), Stem (SAE), and Root (REA) extracts were prepared (50 g/400 mL, shaken for 24 h at 25°C; centrifuged at 2000 rpm/min). The extraction yield was 160, 150, and 240 mL for roots, stems, and leaves, respectively. For each aqueous extract (AE), different concentrations (C1: 25%, C2: 50%, C3: 75%, C4: 100%) were made. C0: 0% was sterile distilled water (control). In the lab, sterilized seeds (20/seeds per Petri dish, 3 replicates) of two okra varieties (Clemson: V1 and Hire: V2) were watered with the different concentrations of LEA, SAE, and REA.
Germination parameters (seed germination percentage (PG), germination index (GI), coefficient of velocity (CVG), mean germination time (MGT), time to 50% germination (T50), latency time (FDG), duration of germination (TSG), last day of germination (LDG), Germination energy (GE: % of germinated seeds after 4 days from the start of the experiment), seed germination index (SGI), mean daily germination (MDG), germination vigor index (GVI), speed emergence (SE), average growth rate (GR), length of the radicle (LR), speed of germination (Spe), inhibition (Inh), phytotoxicity (Tox)) were evaluated [10, 11, 12, 13, 14].
$$\text{Inhibition} = \frac{(\text{control-extract})}{\text{control}} \times 100$$
$$\text{Phytotoxicity} = \frac{1 - (\text{extract}/\text{control})}{100}$$
$$\text{PG}(\%) = \frac{\text{Number of germinated seeds}}{\text{Total seed number}} \times 100$$
$$\text{GI} = \frac{N^0 \text{ of germinated seeds in first count} + N^0 \text{ of germinated seeds in final count}}{N^0 \text{ of germinated seeds in first count} + N^0 \text{ of germinated seeds in final count}}$$
$$\text{CVG} = \frac{(G_1 + G_2 + \cdots + G_n)}{(1 \times G_1 + 2 \times G_2 + \cdots + n \times G_n)} \times 100$$
seeds, n: last day of germination (5)
$$ \mathrm {M G T} = \frac {\sum (D n)}{\sum n}; $$
n ∑ ;With n: N° of seeds germinated on each ∑
day and D: day of counting (6)
( ) ( ) ( ) ( )
0.5 T T G T day T G G
$$ ) = T _ {1} + \frac {\left(T _ {2} - T _ {1}\right) \times \left(0 . 5 - C\right)}{\left(G _ {2} - G _ {1}\right)} $$
; G1: cumulative GR
2 1 1 50 1 2 1
using the value flanking to 50% thru smaller value, G2: cumulative GR using the value flanking to 50% thru greater value, T1: day whose cumulative GR was nearer to 50% by smaller value, T2: day whose cumulative GR was nearer to 50% by greater value (7) GVI = LR (cm) x GI (8) o st o N of germinatedseedson the1 day 100 N of germinatedseedson thelast day × SE = (9) x x x x x y y y $$ - + \frac {x _ {2} - x _ {1}}{v} + \dots \frac {x _ {n} - x _ {n - 1}}{v} $$
1 1 2 1 n n
1 2 n GR=
(10) xn: N° of seeds germinated on day n, yn = N° of days from start of trial to day n MDG = PGF Nj ; PGF: % of final germination, Nj: N° of days to full final germination (11).
SGI=∑Di Ni/W Di: N° of days after seeding, Ni: N° of seeds germinated on day 1, W: whole N° of seeds sowed (12) Spe = [N1/1 + N2/2…]×100; Ni/i: ratio between N° of seeds germinated/day (13)
Data Analysis
The collected data were subjected to analysis of variance using R Studio version 4.0.5 and IBM SPSS Statistics 20 software. Means were compared with each other using Duncan’s test at the 5% threshold. Graphical representations were made using Graphpad software.
Results and Discussion
Seed Germination Percentage
Aqueous extract (AE), variety, and organ of M. pudica significantly (P<0.05) affected the germination of okra seeds. From Day 2, the interaction of variety × treatment × organ was significant (P<0.01). With increasing AE from C1 to C4, a gradual decrease in seed germination parameters was observed in V1 and V2 (Figures 1 & 2, Table 1).
At V1, on Day 9, the highest seed germination percentage was recorded for C0 (Leaf: 68.33±2.89%, Stem: 80±5%, Root: 63.33±2.89%) and the lowest for C4 (Leaf: 1.67±2.89%, Stem: 0%, Root: 8.33±2.89%).
At V2, the highest seed germination percentage was recorded for C0 (Leaf: 81.67±2.89%, Stem: 86.67±2.89%, Root: 63.33±2.89%) and the lowest for C4 (Leaf: 6.67±2.89%, Stem: 3.33±5.77%, Root: 33.33±7.64%).
Toxicity was higher for SAE (Stem AE) and LAE (Leaf AE) compared to RAE (Root AE). V2 showed the best germination percentage.
| SV | PG | PG | PG | PG | PG | PG | PG | PG | PG | MDG | GE | GVI | GR | Spe | SGI | GI | LR | T50 | CVG | MGT |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| day1 | day2 | day3 | day4 | day5 | day6 | day7 | day8 | day9 | ||||||||||||
| Variety (V) | ns | * | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | ns | ns | ns |
| Treatment (T) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Organ (O) | * | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | ns | *** | *** | *** |
| V*T | ns | *** | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | * | ns | ns |
| V*O | ns | * | *** | *** | *** | *** | *** | *** | *** | *** | *** | ns | *** | *** | *** | *** | ns | * | ns | ns |
| T*O | ** | ** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | ** | ** | *** | *** |
| V*T*O | ns | *** | *** | *** | *** | *** | ** | ** | ** | ** | *** | *** | *** | *** | *** | *** | *** | - | ns | ns |
| SV | Inh | Inh | Inh | Inh | Inh | Inh | Inh | Inh | Tox | Tox | Tox | Tox | Tox | Tox | Tox | Tox | TSG | SE | FDG | LDG |
| day2 | day3 | day4 | day5 | day6 | day7 | day8 | day9 | day2 | day3 | day4 | day5 | day6 | day7 | day8 | Day9 | |||||
| Variety (V) | * | ** | *** | *** | *** | *** | *** | *** | * | ** | ns | *** | ns | *** | *** | *** | *** | ns | ns | ** |
| Treatment (T) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | ns | *** | ns | *** | *** | *** | *** | *** | *** | * |
| Organ (O) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | ns | *** | ns | *** | *** | *** | ** | ns | *** | ns |
- V*T ns ns
- ***
- ***
- ***
- **
- **
- ** ns ns ns
- *** ns
- **
- **
- ** ns ns ns ns
- V*O ns ns
- ***
- ***
- * ns ns ns ns ns ns
- *** ns ns ns ns ns ns ns ns
- T*O
- **
- ***
- ***
- ** ns ns ns ns
- **
- *** ns
- ** ns ns ns ns
- *
- **
- ***
- *
- V*T*O
- *
- ***
- ***
- ***
- ***
- **
- **
- **
- *
- *** ns
- *** ns
- **
- **
- ** ns ns ns ns
Table 2: Significance of considered parameters.
SV: Source of variation, PG: germination capacity, GR: Average growth rate, CVG: coefficient of velocity, FDG: latency (first day of germination), GE: germination energy, GI: germination index, GVI: germination vigor index, Inh: Inhibition, LDG: last day of germination, LR: length of radicle, MDG: mean daily germination, MGT: mean germination time, SE: speed emergence, SGI: seed germination index, Spe: speed of germination, T50: time to 50% germination, TSG: duration of germination, Tox: phytotoxicity. *, and *: significant at the 0.05, 0.01 and 0.001 levels, respectively, ns: no significant difference at the 0.05 level.

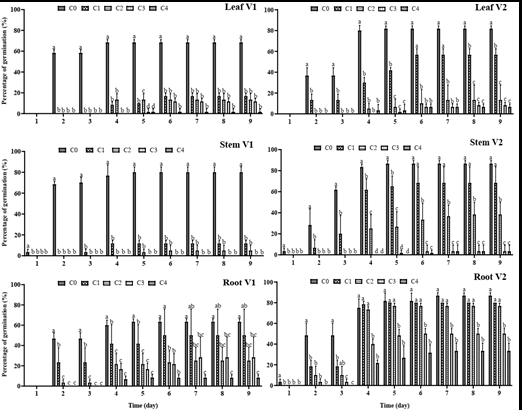
Figure 2: Effect of aqueous extract (AE) on germination percentage of okra’s seed. C0: sterile distilled water (Control), C1: 25% of AE, C2: 50% of AE, C3: 75% of AE, C4: 100% of AE, V1: Clemson, V2: Hire. For each graph, at each time, the average ± standard deviation with the identical inscription was not differing significantly at the 0.05 level of Duncan’s test.
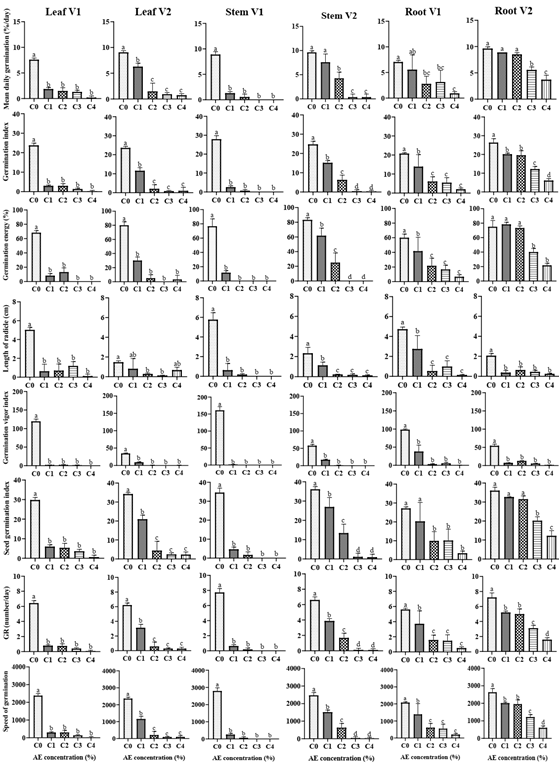
Figure 3: Effect of aqueous extract (AE), organ and variety on germination parameters of okra’s seed. C0: distilled water, C1: 25% of AE, C2: 50% of AE, C3: 75% of AE, C4: 100% of AE, V1: Clemson, V2: Hire. GR: average growth rate. For each graph, the average ± standard deviation with the identical inscription was not differing significantly at the 0.05 level of Duncan’s test.
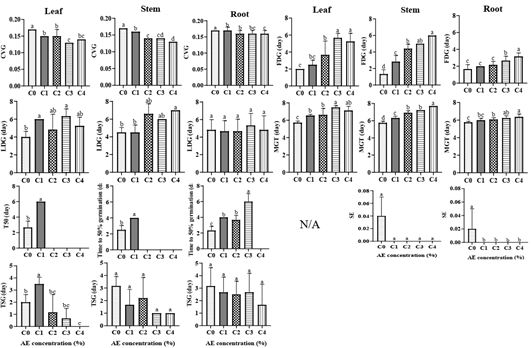
Figure 4: Effect of aqueous extract (AE) and organ on germination parameters of okra’s seed. C0: sterile distilled water (Control), C1: 25% of AE, C2: 50% of AE, C3: 75% of AE, C4: 100% of AE, V1: Clemson, V2: Hire. CVG: coefficient of velocity, FDG: latency (first day of germination), LDG: last day of germination, MGT: mean germination time, SE: speed emergence, T50: time to 50% germination, TSG: duration of germination, N/A: not available. For each graph, the average ± standard deviation with the identical inscription was not differing significantly at the 0.05 level of Duncan’s test.
Speed of Germination
AE, variety, and organ significantly (P<0.001) affected the speed of germination of okra seeds. The interaction between variety × treatment × organ was significant (P<0.001) (Table 1, Figure 3).
With increasing AE from C1 to C4, a gradual decrease in the speed of germination of okra seeds was observed in V1 and V2. V2 showed the highest germination speed with RAE (Figures 5 & 6).
Germination Vigor Index (GVI)
AE, variety, and organ significantly (P<0.001) affected the SGI of okra seeds. The interaction between variety × treatment × organ was significant (P<0.001) (Table 1, Figure 3). AE significantly (P<0.001) reduced the SGI. V2 showed the highest SGI with Root AE.
The lowest SGI was found at C4 (0.58±1.01) for LAE; at C3 (0±0) and C4 (0±0) for SAE; and at C4 (3.38±1.13) for RAE in V1. For V2, the lowest SGI was found at C4 for LAE (2.3±1.39), SAE (0.9±1.56), and RAE (12.25±2.7).
Length of Radicle
AE and variety significantly (P<0.001) affected the length of the radicle. The interaction between variety × treatment × organ was significant (P<0.001) (Table 1, Figure 3). AE significantly (P<0.05) reduced the length of okra radicle. The mean length of radicles observed after the 9th day in V1 Leaf was 5.03, 0.63, 0.7, 1.2 and 0.12 cm respectively for C0, C1, C2, C3 and C4; in V2 Leaf was 1.47, 0.8, 0.28, 0.12 and 0.67 cm respectively for C0, C1, C2, C3 and C4. In V1 Stem, it was 5.77, 0.62, 0.18, 0 and 0 cm respectively for C0, C1, C2, C3 and C4; in V2 Stem, it was 2.33, 1.13, 0.22, 0.18 and 0.13 cm respectively for C0, C1, C2, C3 and C4. In V1 Root, it was 4.73, 2.75, 0.53, 0.98 and 0.13 cm respectively for C0, C1, C2, C3 and C4; in V2 Root it was 2.07, 0.37, 0.63, 0.45 and 0.25 cm respectively for C0, C1, C2, C3 and C4.
Mean Daily Germination (MDG)
AE, variety, and organ significantly (P<0.001) affected the MDG of okra seeds. The interaction between variety × treatment × organ was significant (P<0.001) (Table 1, Figure 3). With increasing AE from C1 to C4, a gradual decrease in the MDG was observed in V1 and V2. V2 showed the highest MDG with RAE (Figures 5 & 6).
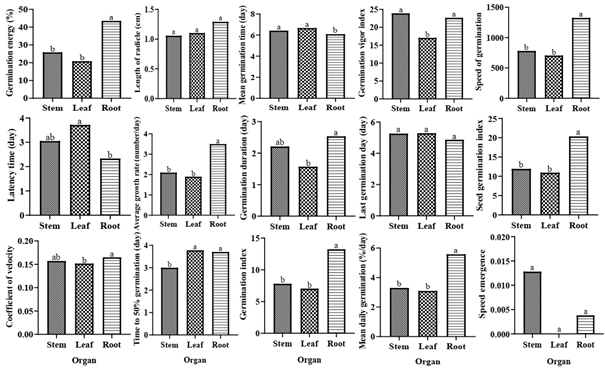
Germination Energy
AE, variety, and organ significantly (P<0.001) affected the GE of okra seeds. The interaction between variety × treatment × organ was significant (P<0.001) (Table 1, Figure 3). With increasing AE from C1 to C4, a continuous decrease in GE was observed in V1 and V2. V2 exhibited the maximum GE with RAE. The lowest GE was observed at C3 and C4 for Leaf AE (0±0); at C1, C2, C3, and C4 for SAE (0±0); and at C4 for RAE (6.67±2.89) in V1. For V2, the lowest GE was observed at C3 for LAE (0±0), at C3 and C4 for SAE (0±0), and for RAE (21.67±2.89).
Germination Index AE, variety, and organ significantly (P<0.001) affected the germination index of okra seeds. The interaction between variety × treatment × organ was significant (P<0.001) (Table 1, Figure 3). With increasing AE from C1 to C4, a continuous decrease in the germination energy was observed in V1 and V2. V2 showed the highest germination energy with RAE (Figures 5 & 6).
Average Growth Rate (GR) AE, variety, and organ significantly (P<0.001) affected the GR of okra seeds. The interaction between variety × treatment × organ was significant (P<0.001) (Table 1, Figure 3). With increasing AE from C1 to C4, a steady decline in GR was noted in V1 and V2. V2 showed the highest GR with RAE.
Latency (FDG)
AE significantly (P<0.001) increased the latency time of okra germination. The variety did not significantly (P>0.05) influence the latency time. Organ significantly (P<0.001) influenced the latency time. The interactions between variety × treatment × organ; variety and treatment; and variety and organ were not significant (P>0.05), but the interaction between treatment and organ was significant (P<0.001).
With increasing AE from C1 to C4, a gradual increase in latency duration was observed (Table 1, Figure 4). The highest latency time was recorded at C3 (5.67±0.52 days) and C4 (5.25±0.96 days) for LAE; at C3 (5.0±0 days) and C4 (6±0 days) for SAE; and at C4 (3.17±0.41 days) for RAE.
Time to 50% germination (T50) The variety did not significantly (P>0.05) influence the T50 of okra seeds. However, AE and organ significantly (P<0.001) influenced the T50. AE and organ (P<0.01), variety and organ (P<0.05), and variety and treatment (P<0.05) interactions were significant. With increasing AE, an increase in T50 was noted (Table 1, Figure 4). T50 was not reached at C2, C3, and C4 for LAE and SAE, and at C4 for RAE.
Mean Germination Time (Mgt)
AE and organ significantly (P<0.001) influenced the MGT of okra. The variety did not significantly (P>0.05) influence this MGT. The interactions variety × treatment × organ, variety × treatment, variety × organ and variety × treatment were not significant (P>0.05), but the interaction treatment × organ was significant (P<0.001).
With increasing AE from C1 to C4, a continuous decrease in MGT was noted (Table 1, Figure 4). The highest value was recorded at C3 (7.48) and C4 (7.13) for LAE; at C4 (7.71) for SAE; and at C4 (6.37) and C3 (6.23) for RAE.
Speed Emergence (SE)
AE significantly (P<0.001) reduced the SE of okra seed. The variety and organ did not significantly (P>0.05) influence the SE. The interactions variety × treatment × organ, variety × treatment, variety × organ were not significant (P>0.05), but the interaction between treatment × organ was significant (P<0.01).
With increasing AE from C1 to C4, a decrease in SE was observed (Table 1, Figure 4). The highest SE was found at C0 for SAE (0.04) and RAE (0.02).
Coefficient of Velocity (Cvg)
AE and organ significantly (P<0.001) influenced the CVG of okra’s seed. The variety did not significantly (P>0.05) influenced CVG. The interactions variety × treatment × organ, variety × treatment, variety × organ and variety × treatment were not significant (P>0.05), but interaction treatment × organ was significant (P<0.001). With increasing AE from C1 to C4, a continuing reducing in CVG was noted (Table 1, Figure 4). The least value was detected at C3 (0.13) and C4 (0.14) for LAE; at C4 (0.13) and C3 (0.14) for SAE; and at C3 (0.16) and C4 (0.16) for RAE.
Duration of Germination (TSG)
AE (P<0.001), variety (P<0.001) and organ (P<0.01) significantly affected the TSG of okra seeds. The interactions variety × treatment × organ, variety × treatment, variety × organ were not significant (P>0.05), but interaction treatment × organ was significant (P<0.05). With increasing AE from C1 to C4, a reducing in TSG was noted (Table 1, Figure 4). The least value was noted at C4 for LAE (0.0), SAE (0.0) and RAE (1.67). V2 showed highest TSG.
Last Day of Germination (LDG)
AE (P<0.05) and variety (P<0.01) significantly affected the LDG of okra seeds. The interactions variety × treatment × organ, variety × treatment, variety × organ were not significant (P>0.05), but interaction treatment × organ was significant (P<0.05). With increasing AE from C1 to C4, a rising in LDG was noted (Table 1, Figure 4). The highest LDG was noted at C3 (6.33±0.82) for LAE, C4 (7±0) for SAE and C3 (5.33±1.37) for RAE. V2 showed highest LDG.
Germination Inhibition (Inh)
AE, variety and organ significantly (P<0.05) affected the germination inhibition of okra seeds. The interaction variety × treatment × organ was significant (P<0.001) (Table 1).
Germination inhibition decreased over time. It increased with increasing AE concentration in the germination medium (Tables 2 and 3). It was highest at C4 concentration (V1C1: 60.64%, V1C2: 78.22%, V1C3: 79.36%, V1C4: 94.8%) (Table 2). Inhibition was higher in the V1 variety than in V2. On Day 9, inhibition was 60.64% in V1C1 versus only 19.94% in V2C1 (Table 2). Inhibition also varied significantly according to the organ from which AE was extracted. It was higher in leaves and stems than in roots. Thus, at Day 10, at C4 concentration, germination inhibition was 94.71% for Leaf, 98.08% for Stem, versus 74.19% for Root (Table 3).
| Var | [AE] (%) | Time (day) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| day 2 | day 3 | day 4 | day 5 | day 6 | day 7 | day 8 | day 9 | ||
| V1 | C1 | 83.45±9.73ab | 74.64±10.41abc | 67.72±10.73bc | 68.19±9.96bc | 60.51±12.23bc | 60.51±12.23bc | 60.51±12.23bc | 60.64±12.16bc |
| C2 | 97.64±1.56a | 87.34± 5.1ab | 81.43±6.13ab | 80.61±5.38ab | 79.04±5.68ab | 78.15±6.24ab | 78.15±6.24ab | 78.22±6.21ab | |
| C3 | 100±0a | 89.9±5.34ab | 90.74±4.9ab | 90.37±4.54ab | 82.82±5.72ab | 79.29±8.49ab | 79.29±8.49ab | 79.36±8.44ab | |
| C4 | 100±0a | 95.96±2.2a | 96.3±2.02a | 94.77±2.28a | 94.77±2.28a | 94.77±2.28a | 94.77±2.28a | 94.8±2.27a | |
| V2 | C1 | 67.32±6.04c | 62.65±7.03c | 27.92±9.98d | 25.64±7.04d | 20.13±4.57d | 20.13±4.57d | 20.13±4.57d | 19.94±4.61d |
| C2 | 93.06±4.59ab | 71.56± 13.46bc | 55.28±14.01Ĉ | 55.91±13.17c | 53.79±11.68c | 51.16±11.1c | 50.52±11.1c | 50.37±11.14c | |
| C3 | 97.68±1.53a | 81.2±9.43abc | 82.22±8.96ab | 79.04±9.6ab | 76.86±8.75ab | 76.86±8.75ab | 76.18±8.61ab | 76.11±8.64ab | |
| C4 | 100±0a | 97.44±1.81a | 88.98±4.71ab | 87.8±5.75ab | 84.52±5.56ab | 83.24±5.76ab | 83.24±5.76ab | 83.19±5.78ab |
Table 3: Inhibition of germination by aqueous extract (AE) of Mimosa pudica (interaction AE*variety). Var: variety, C0: sterile d
Table 2: Inhibition of germination by aqueous extract (AE) of Mimosa pudica (interaction AE*variety). Var: variety, C0: sterile distilled water (Control), C1: 25% of AE, C2: 50% of AE, C3: 75% of AE, C4: 100% of AE, V1: Clemson, V2: Hire. For each day, the average ± standard error with the identical inscription was not differing significantly at the 0.05 level of Duncan’s test.
| Org- an | [AE] (%) | Time (day) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| day 2 | day 3 | day 4 | day 5 | day 6 | day 7 | day 8 | day 9 | ||
| Stem | C1 | 88.1±8.83ab | 81.49±9.54a | 55.28±13.65cd | 55.35±13.8cd | 53.44±15.03bc | 53.44±15.03bc | 53.44±15.03bc | 53.3±15.1bc |
| C2 | 100±0a | 100±0a | 84.94±7.89ab | 82.59±7.37abc | 77.72±8.11ab | 75.8±8.75ab | 74.84±9.27ab | 74.84±9.27ab | |
| C3 | 100±0a | 100±0a | 100±0a | 99.04±0.96a | 98.08±1.92a | 98.08±1.92a | 98.08±1.92a | 98.08±1.92a | |
| C4 | 100±0a | 100±0a | 100±0a | 100±0a | 99.04±0.96a | 98.08±1.92a | 98.08±1.92a | 98.08±1.92a | |
| Leaf | C1 | 81.98±9.01b | 84.49±4.37a | 75.12±5.97abc | 67.24±8.13abc | 53.19±10.2bc | 53.19±10.2bc | 53.19±10.2bc | 53.19±10.2bc |
| C2 | 100±0a | 93.23±1.37a | 87.07±4.04ab | 86.13±4.14ab | 84.1±5a | 82.07±5.1a | 82.07±5.1a | 82.07±5.1a | |
| C3 | 100±0a | 100±0a | 100±0a | 97.76±1.43a | 87.36± 2.47a | 87.36±2.47a | 86.34±2.12a | 86.34±2.12a | |
| C4 | 100±0a | 100±0a | 97.92±2.08a | 96.74±2.15a | 94.71±1.91a | 94.71±1.91a | 94.71±1.91a | 94.71±1.91a | |
| Root | C1 | 56.08±8.89c | 39.98±6.52b | 13.06±11.34e | 18.15±10.47e | 14.34±11.01d | 14.34±11.01d | 14.34±11.01d | 14.37±11.01d |
| C2 | 86.04±5.8ab | 45.11±13.21b | 33.06±14.53de | 36.1±13.9d | 37.42±12.39cd | 36.1±12.14cd | 36.1±12.14cd | 36.04±12.23cd | |
| C3 | 96.53±2.2ab | 56.64±6.54b | 59.44±6.47bcd | 57.3±7.85cd | 54.07±6.86bc | 48.78±8.87bc | 48.78±8.87bc | 48.79±8.86bc | |
| C4 | 100±0a | 90.09±2.6a | 80±4.28abc | 77.13±5.77abc | 75.19±5.78ab | 74.23±6.17ab | 74.23±6.17ab | 74.19±6.21ab | |
| Var | [AE] (%) | Time (day) | |||||||
| day 2 | day 3 | day 4 | day 5 | day 6 | day 7 | day 8 | day 9 | ||
| V1 | C1 | 0.008±0.001b | 0.008±0.001abc | 0.007±0.001bc | 0.007±0.001bc | 0.006±0.001bc | 0.006±0.003bc | 0.006±0.001bc | 0.006±0.001bc |
| C2 | 0.01±0a | 0.009±0.001ab | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001ab | |
| C3 | 0.01±0a | 0.009±0.001ab | 0.009±0.001ab | 0.009±0.001ab | 0.008±0.001ab | 0.008±0.002ab | 0.008±0.001ab | 0.008±0.001ab | |
| C4 | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | |
| V2 | C1 | 0.007±0.001c | 0.006±0.001c | 0.003±0.001d | 0.003±0.001d | 0.002±0.001d | 0.002±0.001d | 0.002±0.001d | 0.002±0.001d |
| C2 | 0.009±0.001ab | 0.007±0.001bc | 0.006±0.001c | 0.006±0.001c | 0.005±0.001c | 0.005±0.001c | 0.005±0.001c | 0.005±0.001c | |
| C3 | 0.01±0a | 0.008±0.001abc | 0.008±0.001abc | 0.008±0.001abc | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001ab | |
| C4 | 0.01±0a | 0.01±0a | 0.009±0.001ab | 0.009±0.001ab | 0.009±0.001ab | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001ab |
Table 4: Inhibition of germination by aqueous extract (AE) of Mimosa pudica (interaction AE*organ). C0: sterile distilled water (
Table 4: Phytotoxicity of aqueous extract (AE) of Mimosa pudica (interaction AE*variety). Var: variety, C0: sterile distilled water (Control), C1: 25% of AE, C2: 50% of AE, C3: 75% of AE, C4: 100% of AE, V1: Clemson, V2: Hire. For each day, the average ± standard error with the identical inscription was not differing significantly at the 0.05 level of Duncan’s test.
| Org- an | [AE] (%) | Time (Day) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| day 2 | day 3 | day 4 | day 5 | day 6 | day 7 | day 8 | day 9 | ||
| Stem | C1 | 0.009±0.001ab | 0.008±0.001a | 0.006±0.001cd | 0.006±0.001cd | 0.005±0.002bc | 0.005±0.002bc | 0.005±0.002bc | 0.005±0.002bc |
| C2 | 0.01±0a | 0.01±0a | 0.009±0.001ab | 0.008±0.001abc | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001ab | |
| C3 | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | |
| C4 | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | |
| Leaf | C1 | 0.008±0.001b | 0.008±0a | 0.008±0.001abc | 0.007±0.001bc | 0.005±0.001bc | 0.005±0.001bc | 0.005±0.001bc | 0.005±0.001bc |
| C2 | 0.01±0a | 0.009±0a | 0.009±0ab | 0.0086±0ab | 0.008±0.001ab | 0.008±0.001ab | 0.008±0.001a | 0.008±0.001a | |
| C3 | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.009±0a | 0.009±0a | 0.009±0a | 0.009±0a | |
| C4 | 0.01±0a | 0.01±0a | 0.01±0a | 0.009±0a | 0.01±0a | 0.01±0a | 0.01±0a | 0.01±0a | |
| Root | C1 | 0.006±0.001c | 0.004±0.001b | 0.001±0.001e | 0.002±0.001e | 0.001±0.001d | 0.001±0.001d | 0.001±0.001d | 0.001±0.001d |
| C2 | 0.009±0.001ab | 0.005±0.001b | 0.003±0.001de | 0.004±0.001de | 0.004±0.001cd | 0.004±0.001cd | 0.004±0.001cd | 0.004±0.001cd | |
| C3 | 0.009± 0ab | 0.006±0.001b | 0.006±0.001cd | 0.006±0.001cd | 0.005±0.001bc | 0.005±0.001bc | 0.005±0.001bc | 0.005±0.001bc | |
| C4 | 0.01±0a | 0.009±0.001a | 0.008±0.001abc | 0.008±0.001abc | 0.008±0.001ab | 0.007±0.001ab | 0.007±0.001ab | 0.007±0.001ab |
Table 5: Phytotoxicity of aqueous extract (AE) of Mimosa pudica (interaction AE*organ). C0: sterile distilled water (Control), C1
Phytotoxicity (Tox)
AE, variety and organ significantly (P<0.05) affected the phytotoxicity of Mimosa pudica AE on germination of okra seeds. Interaction variety × treatment × organ was significant (P<0.001) (Table 1).
Overall, the phytotoxicity of M. pudica AEs on okra seed germination decreased over time. It increased with increasing AE concentration in the germination medium (Tables 2 & 3). It was highest at C4 concentration (C1_Root: 0.001, C2_Root: 0.004, C3_Root: 0.005, C4_Root: 0.007) (Table 5). Phytotoxicity was higher in the V1 variety than in V2. On Day 9, it was 0.006 for V1C1 versus only 0.002 for V2C1 (Table 2).
Phytotoxicity also varied significantly according to the organ from which the AE was extracted. Thus, it was higher in leaves and stems than in roots. At Day 10, from C1 to C4, AE phytotoxicity ranged from 0.005 to 0.01 for LAE, from 0.005 to 0.01 for SAE, and from 0.001 to 0.007 for RAE (Table 3) [10, 11, 12, 13].
Discussion
With increasing AE from C1 to C4, an increase in latency duration, mean germination time, time to 50% germination, germination inhibition, and phytotoxicity was observed. Similar results were reported by Pratap Chandran R [12] on Brassica juncea. With increasing AE from C1 to C4, a decrease in the percentage of germination, speed of germination, seed germination index, germination vigor index, length of radicle, germination energy, germination index, mean daily germination, coefficient of velocity, speed of emergence, duration of germination, and last day of germination of okra seeds was observed. Similar results were reported by Pratap Chandran R [12] on Brassica juncea, [14] on Poaceae, and [15] on Bidens pilosa.
The different AEs of M. pudica negatively affected all the germination parameters evaluated. The negative impact of M. pudica AEs on okra seed germination is attributed to the action of allelochemicals in the extracts. Indeed, they negatively affect the physiological mechanisms throughout germination. Mimosine is a key allelochemical present in the Mimosa genus, reported to be responsible for the high allelopathic action by inhibiting the germination of crop seeds, and it is found in great amounts in the leaves, pods, and seeds of tropical pulses [9, 12]. The reduction in radicle length by AEs may be caused by the inhibition of cell division and cell elongation.
AE toxicity was higher in leaves and stems than in roots. This result is similar to those of this result could be explained by the higher levels of allelochemicals in the leaves and stems of M. pudica. It could also be explained by the different chemical compositions of the tested organs, hence the different allelopathic effects [13, 14, 15]. Burial is one of the most popular agricultural practices in the region, aiming to restore soil fertility. However, given the presence of very high allelochemical substances in the leaves and stems that inhibit the germination of okra seeds, the removal of the residues of this weed from the plots should be encouraged to increase the rate of seedling emergence and consequently the yield of okra in the locality.
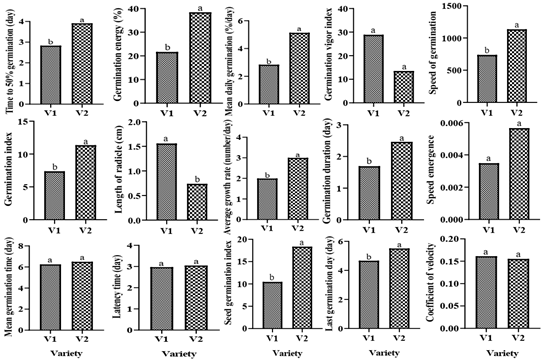
Variety V2 was less affected by AEs compared to V1. Variety inconsistency related to germination traits may be explained by the heterogeneity of their genetic make-up [16, 17]. Indeed, due to its intrinsic characteristics, a variety can be more resistant to diseases [18, 19], more suitable for symbiosis [20, 21, 22, 23, 24, 25], or more tolerant to stress than another [26, 27, 28, 29]. These traits thus give it a better adaptation to the different hazards of the environment.
The phytotoxic effect of Mimosa species causes root growth retardation in target weeds such as Echinochloa crus-galli (barnyard grass) and Ruellia tuberosa L. [30, 31, 32]. It could also be tested on local target weeds competing with okra, such as Commelina benghalensis L., Echinochloa pyramidalis (Lam.) Hitchc. & Chase, Bidens pilosa L., etc. The extraction costs and application methods of Mimosa species as bioherbicides have not yet been documented. Conventional (sprays, granules) and modern (application platforms with sensors, drones, etc.) application methods can be tested.
Conclusion
This work reports the inhibitory effect of M. pudica on all okra seed germination parameters depending on the concentration of root, stem or leaf extracts. The seeds of V2 were less affected by the extracts compared to those of V1; the V2 variety is therefore recommendable to farmers in this region. Inhibition was significantly stronger with leaf and stem aqueous extract compared to the root. The burial of this weed is therefore not recommended for better emergence of okra seeds. In view of its allelopathic effect on okra seed germination, our findings suggest that M. pudica leaf and stem extracts should be tested on weeds to ensure their bioactivity as they can be utilized as a source of natural herbicides.
References
-
Kwok CTK, Ng YF, Chan HTL, Chan SW (2025) An overview of the current scientific evidence on the biological properties of _Abelmoschus esculentus_ (L.) Moench (Okra). Foods14 (2): 177.
-
Siemonsma JS, Kouamé C (2004) _Abelmoschus esculentus_ (L.) Moench. In: Grubben GJH, Denton OA (Eds.), PROTA (Plant Resources of Tropical Africa), Wageningen.
-
Mohammed HY, Teferra TF, Sime G (2024) Indigenous knowledge and nutritional and morphological characterization of okra (_Abelmoschus esculentus_ (L.) Moench) varieties in Western Ethiopia. Food Sci Nutr 12(4): 2537-2550.
-
Singh AK, Abrol GS, Mishra V, Singh RK (2023) Nutritional, antioxidant properties and health benefits of Okra. In: Mani A, et al. (Eds) Antioxidant properties and health benefits of horticultural crops. Brillion Publishing, New Delhi, India, pp: 377-395.
-
FAOSTAT (2024) Statistics. FAO, Rome, Italy.
-
Ngale TUP, Lombeko OTV, Suh C, Temegne NC, Chimi NLL, et al. (2023) New fertilization approach improves okra (_Abelmoschus esculentus_ L. Moench) production on acidic and degraded soil in Cameroon. Eur J Agric Food Sci 5(2): 13-22.
-
Fomekong MK, Tetang EFT, Temegne CN, Atabong PA, Ntsomboh GN, et al. (2024) Effect of halopriming on seed germination performance and early seedling establishment in okra (_Abelmoschus esculentus_ (L.) Moench) under saline conditions. Plant Physiol Rep 30: 146-159.
-
Chopra N, Tewari G, Tewari LM, Upreti B, Pandey N (2017) Allelopathic effect of _Echinochloa colona_ L. and _Cyperus iria_ L. weed extracts on the seed germination and seedling growth of rice and soyabean. Adv Agric 1-5.
-
Gunawardhana CB, Shakkya J, Ranasinghe, Waisundara VY (2015) Review: _Mimosa pudica_ Linn: the garden weeds with therapeutic properties. Int J Plant Sci 62(4): 234-241.
-
Temegne NC, Liégui GS, Sandeu KDB, Fomekong KM, Mbadia KNS, et al. (2024) Manure-based composts influence soil quality after lettuce (_Lactuca sativa_ L.) production. J Saudi Soc Agric Sci.
-
Ashti SA, Hero FHK, Dlshad AO, Nawroz AT (2018) Response of some plant species towards the allelopathy of two types of chickpea (_Cicer arietinum_ L.) seed extracts. Appl Ecol Environ Res 16(6): 8119-8129.
-
Pratap Chandran R, Deepak V, Krishna S, Fathima S, Ameena T, et al. (2018) Allelopathic activity of leaf extracts of _Mimosa pudica_ on growth parameters of _Brassica juncea_ seeds. BAOJ Biotech 4: 033.
-
Tahir NA, Azeez HA, Hama Amin HH, Rashid JS, Omer DA (2019) Antibacterial activity and allelopathic effects of extracts from leaf, stem and bark of Mt. Atlas mastic tree (_Pistacia atlantica_ subsp. kurdica) on crops and weeds. Allelopathy J 46(1): 121-132.
-
Souza FA, Mourão JrM (2010) Response pattern of _Mimosa pudica_ e _Senna obtusifolia_ to potentially allelopathic activity of Poaceae species. Planta daninha 28(spe): 927-938.
-
Wang Jh, Shi Sj, Chen W, Li Y, Cui Xl (2020) Allelopathy and invasiveness of _Bidens pilosa_ and _Mimosa pudica_. Acta Prataculturae Sinica 29(4): 81-91.
-
Temegne NC, Mbogne TJ, Nbendah P, Youmbi E, Taffouo VD, et al. (2015) Effect of phosphate deficiency on growth and phosphorus content of three Voandzou (_Vigna_ _subterranea_ (L.) Verdc.) varieties. IOSR-JAVS 8(9): 52-59.
-
Mouafor BI, Temegne NC, Ngome AF, Malaa D (2016) Farmer’s adoption of improved cassava varieties in the humid forest agro-ecological zone of Cameroon. Greener J Agric Sci 6(10): 276-284.
-
Tsala R, Ngatsi PZ, Temegne NC, Dida LSL, Kuaté TWN, et al. (2022) Efficacy of aqueous extract of the seeds of _Annona_ _muricata_ L. in the control of late blight (_Phytophthora_ _infestans_) of tomato (_Lycopersicon esculentum_ Mill.) in the field. J Mod Agric Biotechnol 1(1): 6.
-
Ngouana LST, Tonfack LB, Temegne CN, Agendia AP, Youmbi E (2023) Current status of strawberry (_Fragaria_ spp.) cultivation and marketing in Cameroon. J Agric Food Res 14 (2023): 100761.
-
Kamtchoum SM, Nchoutnji I, Temegne CN, Tonfack LB, Fofe L, et al. (2019) Comparative effect of biological fixation of nitrogen and chemical fertilizer on yield optimization of two sorghum varieties in the Western Highlands. Asian J Agric Hortic Res 4(3): 01-10.
-
Temegne NC, Nkou Foh TD, Taffouo VD, Wakem GA, Youmbi E (2018) Effect of mycorrhization and soluble phosphate on growth and phosphorus supply of Voandzou (_Vigna subterranea_ (L.) Verdc.). Legume Res 41(6): 879-884.
-
Maffo AF, Ngonkeu ELM, Chaintreuil C, Temegne CN, Ntsomboh Ntsefong G, et al. (2022) Morphological and molecular diversity of arbuscular mycorrhizal fungi associated to _Carica papaya_ L. rhizosphere in two agro- ecological zones in Cameroon. Afr J Agric Res 18(8): 632- 646.
-
Temegne NC, Nkou Foh TD, Taffouo VD, Ntsomboh Ntsefong G, Youmbi E (2017) Influence of mycorrhization and phosphate fertilizer on growth of Voandzou (_Vigna_ _subterranea_ (L.) Verdc.). Int J Biol Chem Sci 11(6): 2587- 2593.
-
Temegne NC, Wakem GA, Taffouo VD, Mbogne TJ, Onguene AN, et al. (2017) Effect of phosphorus fertilization on arbuscular mycorrhizal fungi in the Bambara groundnut rhizosphere. Afr J Microbiol Res 11(37): 1399-1410.
-
Temegne NC, Gouertoumbo WF, Nkou Foh TD, Wakem GA, Ntsefong NG, et al. (2019) Effect of single superphosphate and arbuscular mycorrhizal fungi on growth and Bambara groundnut (_Vigna subterranea_ (L.) Verdc.) yield. KFokam J App Sci Technol 1(Inaugural Edition): 4-13.
-
Temegne NC, Gouertoumbo WF, Nana AS, Taffouo VD, Youmbi E (2024) Effect of phosphorus fertilizer on morpho-physiological, and biochemical traits of Bambara groundnut (_Vigna subterranea_ (L.) Verdc.) plants under water deficit. Asian J Res Crop Sci 9(2): 142-155.
-
Temegne NC, Taffouo VD, Tadoh TC, Gouertoumbo WF, Wakem GA, et al. (2019) Effect of phosphate fertilization on growth, yield and seed phosphorus content of Bambara pea (_Vigna subterranea_) landraces. J Anim Plant Sci 29(3): 703-713.
-
Temegne NC (2018) Improvement in the performances of Voandzou (_Vigna subterranea_ (L.) Verdc.) in response to phosphate deficiency through chemical and biological fertilization. Ph.D Thesis, University of Yaounde I, Faculty of Science, pp: 51.
-
Kamtchoum SM, Nuemsi PPK, Tonfack LB, Edinguele DGM, Kouahou WN, et al. (2018) Production of Bean (_Phaseolus vulgaris_ L.) under organo-mineral fertilization in humid forest agro-ecological zone with bimodal rainfall pattern in Cameroon. Annu Res Rev Biol 29(4): 01-11.
-
Koodkaew I, Senaphan C, Sengseang N, Suwanwong S (2018) Characterization of phytochemical profile and phytotoxic activity of _Mimosa pigra_ L. Agric Nat Resour 52: 162-168.
-
Khang DT, Quy TN, Dam NP, Tuan NT, Men TT, et al. (2023) Isolation and purification of potential weed inhibitors from _Mimosa pigra_ L. Heliyon 9(7): e18205.
-
Koodkaew I, Wannathong R (2018) Effects of _Mimosa_ _pigra_ L. leaf extract on growth behavior of _Ruellia_ _tuberosa_ L. and Echinochloa crus-galli (L.) P. Beauv. APST 23(1).
- Community Forestry Enterprises as a Model for Sustainable Forest Development: The Case Of The "Baja Tarahumara" in Chihuahua, Mexico
- Ecological and Socio-Economic Impacts of Chromolaena odorata and Mesosphaerum suaveolens, Two Invasive Alien Species in Central and Southern Benin, West Africa
- Epigenetic Sustainability: Modeling the Human Factor as a Natural Resource through Science 4.0 and the NR3C1 Biological Pilot
- Growth-at-Risk: A Framework for Assessing Economic Vulnerability
- The Rural Territory as a Socioecological System for the Management of Public Policy for Sustainable Rural Development
- Transition Risks in a Small Open Economy: An EDSGE Model with Blue Firms, Financial Frictions and Macroprudential Policy