Postharvest Control of Strawberry Fruit Soft Rot by Laminaria Digitata Extracts
The aim of this work was to assess the potential suppression of extracts of Laminaria digitata for controlling postharvest soft rot of strawberries by Rhizopus stolonifer. The antifungal activity of raw extract and fractionated by hexane, ethanol and water was in vitro measured at a concentration range. The activity of the raw extract was in vivo assessed at the same concentration range in comparison to fungicide Fenhexamid. Observations by scanning electron microscopy of R. stolonifer were performed. The peroxidase activity in L. digitata-treated inoculated strawberries was assessed. A significant inhibition of mycelia growth (until 80% after 5 days of incubation) and suppression of sporangia germination (up to 95% after 24 hours) were found applying 30 g L-1 raw extract. Only extracts fractionated by hexane and ethanol were suppressive against mycelia (until 71% and 67% respectively) and sporangia (up to 82% and 69% respectively). Fruit decay inhibition on the R. stolonifer/strawberries pathosystem increased up to 75% after 3 days of incubation in preventive treatment after application of increasing doses of the extracts. Efficacy of the treatments was related to dose of extract in all the trials. The increased peroxidase activity (up to 7.04 ΔOD420 g−1 min−1) observed in fruit tissue after one-day from application of 30 g L-1 raw extract suggests that suppression could also be due to activation of systemic acquired resistance in artificially infected strawberry fruit.
Introduction
Strawberry (Fragaria × ananassa Duch.) fruit is one of the most popular and widely consumed fresh o minimally processed fruit worldwide that requires massive use of pesticides [1, 2]. It is a very perishable commodity due to industrial processing chains that occurs immediately after harvesting which cause mechanical injury, desiccation, physiological disorders, deterioration of quality and nutrient composition, decay, abiotic stress, mycotoxin Postharvest Control of Strawberry Fruit Soft Rot by Laminaria Digitata Extracts contamination, and reduction of their market value [3, 4]. The main causes of strawberry fruit decay in the retail points are due to development of rots caused by spoilage fungi [5]. The postharvest soft rot of strawberry fruit caused by several species of Rhizopus or Mucor can be considered as the most devastating in the retail points, as well as the relative etiological fungi can spread rapidly from infected fruit to the healthy fruit which can result in extensive J Agri Res
disease breakdown [5]. Among the most important postharvest diseases in strawberry fruit, soft rot caused by the zygomycete fungus Rhizopus stolonifer (Ehrenb.: Fr.) Vuill. Induces economically severe loss both in the field and overall during long-distance transport, especially if surface of strawberries is mechanically damaged during handling and transportation and when storage temperature is more than 4–5°C [6, 7]. R. stolonifer is considered as one of the fastest-growing fungi, particularly under higher moisture condition, being considered one of the most devastating postharvest spoilage funguses since it is considered highly invasive showing faster development of aerial mycelia under common storage temperature [8]. R. stolonifer colonizes host tissue at lower temperatures, usually from 5 to 10°C, than other spoilage fungi, such as Botrytis cinerea Pers. ex Fr. which needs more than 15°C for fruit colonization. Moreover, it has a wide array of hosts that make it one of the most destructive of fungi [9]. Although control of strawberry fruit postharvest decay could be achieved by physical, chemical and biological methods, nevertheless only protective broad-spectrum fungicides, or their mixtures, can be used to effectively control soft rot during the fruit-ripening stage [10, 11, 12, 13]. R. stolonifer has become an important target of control by several synthetic fungicides as Boscalid, Fludioxinol, and Fenhexamid due to its wide array of hosts and fast colonization [5]. Although the appearance of resistant strains and the concern for residues on fruit and in the environment focused producers to explore some alternatives to fungicides as hydrocooling, hot water dips, waxes and biocontrol agents, any satisfying solution nevertheless has still been found [14]. In addition, the requirements from consumers for fruit-free by chemical residues have addressed researches to find safer alternatives for environmental and human health based on a wide range of natural antifungal compounds [15]. For example, phenolic substances given from wild edible herbs and terpenic compounds provided from aromatic plants are considered among the best candidates for controlling many fruit spoilage fungi [16, 17, 18]. Very fewer papers on the alternative strategies for controlling postharvest strawberries soft rot without use of chemical fungicides were reported. Authors have tested the antifungal efficacy of essential oils in controlling postharvest decay of strawberry fruit by R. stolonifer in place of fungicides [19]. Others authors have assessed the activity of calcium chloride and lemongrass oil, alone or in combination, against R. stolonifer on peaches under postharvest condition as alternative to fungicides, demonstrating that the these two treatments gave the best performance against peaches soft rot and that the control effect was synergic for disease severity [20]. Seaweeds are nowadays considered as a new source of natural bioactive substances to stimulate growth in plants by protecting them from pathogens, physiological hazards and several physiological disorders under storage condition. Seaweed is usually employed as a liquid extract through soil irrigation, or spray treatment onto the leaves of tomato, apple, wheat, strawberry, and winter rapeseed [21, 22]. Besides their use as organic fertilizers and plant growth promoters, seaweed extract can enhance the tolerance or resistance of crops against a wide range of abiotic and biotic stresses extending the postharvest shelf life of fruit [23, 24, 25, 26]. Moreover, seaweed extract have a great potential as disease crop protective, either through a direct effect against fungal plant pathogens or indirectly through induction of resistance in plants by enhancing the activities of various defense-related enzymes that can help the host tissue to limit fungal colonization [27, 28, 29, 30]. Raw extracts from brown seaweeds contain a wide range of antifungal substances mainly belonging to lipids (triglycerides and fatty acids), phenolic compounds (phlorotannins), and water-soluble polysaccharides (laminarans, fucoidans and alginates) which have been characterized and investigated for the antifungal properties [31]. In previous study, the use of crude extracts from a collection of brown and red seaweeds in preventing fruit postharvest losses caused by several plant pathogenic fungi has been evaluated [32]. The aim of this work was to assess the potential use of four extracts from the brown seaweed Laminaria digitata (Huds.) Lamouroux for effective controlling postharvest soft rot by R. stolonifer of strawberries. The first objective was to investigate the potential suppression of L. digitata raw extract in preventing soft rot under in vitro and in vivo conditions. The second one was to clarify the most probable mechanisms of action explaining the suppressive effect observed.
Materials and Methods
L. digitata Extracts
About 100 Kg of fresh algal biomass harvested during 2017 from healthy and matured L. digitata cultures performed into a photo-bioreactor of 1,000 L capacity (Braun Biotech International BIOSTAT D-1000, Milan, Italy), was purchased from a marine biorefinery located on the coastal areas near to Agadir beach site (Morocco). Algal biomass was immediately refrigerated after harvesting, thoroughly washed with seawater and washed with tap water to remove all extraneous particles and epiphyte organisms. Biomass was dried into an industrial spray drier located at ENEA – Trisaia Research Centre (Policoro, Matera, Italy) – chopped, finely pulverized, heat-treated for 24 h with sodium hydroxide (1:10 w/w) to triglycerides saponification and stored at 4°C until extraction of potentially bioactive substances [22]. Extraction was carried out for three time (from 4 h to 5 h for each step) using a mixture of un-polar and polar solvents in appropriate proportions (n- hexane/ethanol/water, 1:2:2 v/v) to extract a wider range of bioactive substances accordingly to method described by Kulandaivel S [33]. Raw extract was pooled, suspended in n-hexane/ethanol/water and stored into glass bottles at 4°C in the dark. Stocks of extract were completely dried under reduced pressure at 35°C using a rotary evaporator (Strike 202, Steroglass, Perugia, Italy) equipped with a vacuum membrane pump (Vacuubrand GMBH, Germany) to eliminate the possibility of inhibitory residues of solvent in the final solution, that will become tested for its potential suppressive property and stored into Erlenmayer flasks at −20°C in the dark till further uses. Aliquots of 50 g raw dry extract were fractionated into three fractions using a range of solvents with different affinity towards fatty acids (n-hexane), phenolic compounds (ethanol), and water-soluble polysaccharides (water) [34]. Dried samples were collected from each flask by a spatula and suspended in n-hexane, or 100% ethanol, or distilled water (1:5 w/v) in separating funnel for 20 days at room temperature to perform extraction of the bioactive substances. Each suspension was individually filtered using Whatman filter paper and re- dried by a rotary evaporator until extract become as a syrup. Three extract fractions were separated from this residue, individually collected from the respective funnel and stored into glass tubes at −20°C in the dark until assayed.
Pathogen
One strain of R. stolonifer (ENEA str. 43243) isolated from a significant sample of diseased strawberry fruit showing soft rot symptom and randomly collected from a storage commercial factory located at Policoro area, was used in all trials. It was isolated from 60 naturally infected fruit and identified basing on the morphological and biochemical characteristics as described by Lima JN, et al., and deposited in the fungal collection of the Laboratory of Mycology and Plant Diseases of ENEA – Trisaia Research Centre [35]. The isolate was maintained under its mono- conidial form on potato dextrose agar (PDA, Sigma- Aldrich, Milan, Italy) slants at 4°C until used for bioassays.
In vitro Antifungal Effects
The antifungal activity of four extracts of L. digitata (raw and fractionated by hexane, ethanol, and water) was in vitro measured vs. pure cultures of R. stolonifer which affects postharvest soft rot of strawberry fruit. Stocks of each dry extract were collected by a sterile spatula, suspended in sterile 0.1 M K-phosphate buffer and tested at the concentration range of 10 g L−1, 20 g L−1 and 30 g L−1 for determining minimum bioactive concentration by poison food technique [36]. Mycelia inhibition was quantitatively assessed by measuring radial growth in Petri plates (100 mm diameter) containing PDA adding 18 mL PDA per plate. In treated plates, aliquots of 2 mL sterile stock suspension containing extracts were added to 18 mL PDA at 42 ± 3°C before solidification. In untreated plates used as control, 2 mL of sterile buffer was added to 18 mL PDA in place of the extract. Three mycelia plugs measuring 5 mm diameter each were cut out by the margin of one-day-old fungal cultures actively growing and aseptically placed on the upper PDA surface. Treated and control plates were incubated in the dark at 18 ± 1°C. Mycelia growth inhibition (MGI%) was measured after 1, 3 and 5 days of incubation with respect to the control plates by the index:
MGI% = [(Dco − Dse) / Dco] × 100;
(Dco = average of colony diameter in the control plates; Dse = average of colony diameter in the amended plates). All experiments were carried out with three replications of ten plates for each. Sporangia germination was evaluated on micro- cultures performed by a microassay on glass slides that allowed the quantitative analysis of sporangia suppression using the optical microscopy technique [16, 17]. Assays on 96-microwell (100 μL by volume) plates purchased from AES Laboratory (Milan, Italy) were performed. Each micro-well was set up with three replicates containing 10 μL potato dextrose broth (PDB, Sigma-Aldrich), 2 μL cellular suspension containing 108 CFU mL-1, and 88 μL of the extracts suspended in sterile 0.1 M K-phosphate buffer for testing sporangia suppression at the concentration range of 10, 20 and 30 g L−1. One micro-well row filled with 10 μL PDB, 2 μL cellular suspension, and 88 μL buffer alone was used as control. Each plate was incubated at 18 ± 1°C. Aliquots of 5 μL sporangia cultures taken from each micro-well were sampled and mounted on the upper surface of glass slides sterilized with denatured ethanol. The number of the total, un-germinated and collapsed sporangia was estimated by a Burker’s hemocytometer using a photomicroscope (40 × magnification) (BX60, Olympus, Milan, Italy). Sporangia germination suppression (SG%) was measured after 2, 10, 18 and 24 hours of incubation by the ratio:
SG% = Su/St × 100;
(Su = average of number of un-germinated and/or collapsed sporangia in cultures supplemented with extract, including control cultures; St = average of number of total sporangia in the same sample). All measurements were performed with three replications of five glass slides taken from each micro-well.
Scanning Electron Microscopy
R. stolonifer strain was grown at 18 ± 1°C in PDB amended with 20 g L-1 L. digitata raw extract for 3 days. PDB supplemented with sterile buffer without extract served as control. Fungal materials were collected and fixed in 2.5% glutaraldehyde at 4°C for 24 h and post- fixed in 1% osmium tetraoxide for 1 h at room temperature [37]. They were dehydrated with ascending concentrations of acetone, critical point dried, and sputter coated with gold. Samples were observed by the scanning electron microscopy (SEM) technique, and observations were carried out on three independent replicates of five aliquots for each by a SEM microscope JSM-1200 EX (JEOL, MA, USA). Samples were collected with a sterile forceps and dried to eliminate water to avoid cell collapse when the surface of it was exposed to high vacuum. Seventy-nm-thickness sections were obtained using an ultra-microtome Power Tome-X (RMC Products, USA) fitted with a glass knife and placed directly onto quartz cuvettes. Sections were mounted in a coater JEOL JSC 1200 (USA) and coated with a thin layer of gold such preventing the build-up of an electrical charge on surface and giving a better image.
In vivo Antifungal Effect
The antifungal activity of raw extract was in vivo measured on inoculated strawberries under postharvest condition in comparison to Fenhexamid. Preventive and curative treatments were both evaluated on strawberries cv. Camarosa harvested from growers located in Basilicata (Policoro) and grown under glasshouse condition (tunnel). Healthy fruit untreated with synthetic fungicides and selected for uniform size, colour, same ripening stage and absence of visible defects and injuries, were washed twice under running tap water for 5 min, surface-disinfected by dipping in 2% sodium hypochlorite solution for 1 min, rinsed with tap water and allowed to air dry. Fruit were injured along the equatorial axis in two opposite points at fixed dimensions (wide = 1 mm, deep = 1 mm), treated with the raw seaweed extract and inoculated with a sporangial suspension of R. stolonifer. Trials treated with the stocks containing 10 g L-1, 20 g L-1 and 30 g L-1 raw extract suspended in sterile 0.1 M K- phosphate buffer were set up. Two controls replacing the extracts, the first one with the buffer alone and the second one with 1.2 g L−1 Fenhexamid-50% as active substances, were both included in the assays during preventive and curative treatments. Aliquots of 30 μL of the stock suspension of extract were dispensed over each wound of the treated fruit allowing the droplet to be absorbed into the fruit. Aliquots of 30 μL of sterile buffer alone, or fungicide, were dispensed over the wound of the control fruit. Each wound was inoculated with 10 μL of a sporangial suspension containing 106 CFU mL−1. For enhancing plant defenses before inoculation, in preventive treatments the pathogen was inoculated over the injured area two days later from application of the extract, or the fungicide, or the buffer alone. Instead, in those curative, the extract, or the fungicide, or the buffer alone, were applied over the wound eight hours later from inoculation of the pathogen to allow sporangia germination before treatment. All fruit were placed in trays, packaged in plastic bags and maintained into a climatic room at 18 ± 2°C and 96 ± 2% RH in the dark for three days. Trials were arranged in a completely randomized experimental design including six replicates per treatment. Thirty fruit per species with two wounds per fruit were considered per each replication. All experiments were repeated twice in two consecutive seasons. Disease incidence was assessed by counting the number of the infected wounds on each fruit, and data were converted into the fruit decay inhibition data (DI%) by the index:
DI% = [(Nco − Nse) / Nco] × 100 (Nco = average of number of infected wounds in the control plot treated with the buffer alone; Nse = average of number of infected wounds in the plots treated with the extracts or with Fenhexamid).
Peroxidase Activity
In order to assess the peroxidase (POD) activity, trials were arranged by thirty Camarosa strawberry fruit previously treated with 30 g L−1 L. digitata raw extract and inoculated with R. stolonifer at the same conditions afore-described. Fruit were sampled and analysed after one day and three days of incubation at 18 ± 2°C and of 96 ± 2% RH in the dark. Inoculated and healthy fruit treated with sterile 0.1 M K-phosphate buffer without extract were both included as positive and negative controls, respectively. Small pieces of fresh tissue (diameter = 2–3 mm, deep = 3–4 mm) were randomly collected from each fruit in four points along the equatorial axis, weighed and powdered with liquid nitrogen. Samples of dry weigh tissue (1 g) for each trial were extracted with 2 mL 0.1 M sodium-phosphate buffer (pH 7.0) at 4°C, suspended in 5 mL sterile water and incubated in a rotary shaker (140 rpm) at 25°C for 2 h. After incubation, the suspension was centrifuged (16,000_g_) at 20°C for 5 min. The supernatant was collected, filtered through a 0.22-μm membrane filter and concentrated to one-tenth of the initial volume in a rotary evaporator at 40°C equipped with a vacuum membrane pump. The concentrated sample was stored at –20°C with 20% glycerol until tested. Each filtrate was used to determine POD activity as described by Hammerschmidt R, et al. with some modifications [38]. The reaction mixture consisting of 2 mL filtrate (each derived from one gram of dry weigh tissue), 1.5 mL of 0.05 M pyrogallol, 0.5 mL of 1.5 Units mL−1 enzyme, and 0.5 mL of 1% H2O2 was incubated at 28 °C for 3 min. Absorbance of the samples, measured at a wavelength of 420 nm, was recorded every 30 sec, and the boiled enzyme preparation served as blank. The POD activity was expressed as change in absorbance of the reaction mixture on a fresh weigh basis (ΔOD420 g−1 min−1).
Data Analysis
The averaged data of mycelia growth inhibition, sporangia germination suppression, and fruit decay inhibition were compared either among the three seaweed-extract doses or the different incubation times (for MGI% and SG%), and either between the three extract doses or the two treatment types (for DI%), by applying two-way variance analysis (two-way ANOVA). Percentage data were transformed into Bliss angular values (arcsin √%) to normalize their distribution before analysis, however the values are shown as untransformed values. In addition, the averaged data regarding to decay inhibition index obtained from the two experiments carried out in different seasons were pooled and treated as a mean alone whenever a preliminary statistical analysis performed with the Bartlett’s test determined homogeneity of variances without significant interactions among them. The averaged data of POD activity were instead compared among the three experimental plots at the same sampling time by using one-way ANOVA. All data were analyzed by the Duncan’s multiple range test (DMRT) at a probability P ≤ 0.05 level whenever ANOVA revealed significant differences among the means. Statistics were carried out using the software pack SPSS programme, v. 12.0 Statistics Base™, Chicago, Illinois, USA.
Results
In vitro Effects
The data set on the effects of different concentrations of L. digitata raw extract and fractionated by hexane, ethanol and water on MGI% and SG% of R. stolonifer at the different incubation times are reported in the tables 1 and 2, respectively. In general, mycelia growth reduction and sporangia germination suppression increased either with the concentration of extract or with the incubation time after supplementation with both raw extract and fractionated extracts by hexane and ethanol. While, any suppressive effect was observed after amendment with extract fractionated by water. In particular, a significant mycelia growth inhibition (80% after 5 days of incubation) and sporangia germination suppression (95% after 24 hours) were found applying 30 g L-1 raw extract. While, the extracts fractionated by hexane and ethanol resulted suppressive to mycelia development until 71 and 67%, respectively, and sporangia germination up to 82 and 69%, respectively, by testing the same extract dose at the end of incubation time. No inhibitor effect applying the buffer without extract was found. Mycelia growth inhibition index and sporangia germination suppression ratio were both significantly affected by the raw extract concentration added into growing media and incubation time (Table 3) because significant interactions among these two factors were shown (P-value < 0.05).
| Concentration of extract (g L-1) | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10 | 20 | 30 | |||||||||||||||||||||||||
| Extract | |||||||||||||||||||||||||||
| Incubation time (days) | |||||||||||||||||||||||||||
| 1 | 3 | 5 | 1 | 3 | 5 | 1 | 3 | 5 | |||||||||||||||||||
| Raw | 11a±0.21a | 28b±0.31 | 32c±0.36 | 38c±0.36 | 47d±0.43 | 59e±0.53 | 45d±0.43 | 61e±0.58 | 80f±0.75 | ||||||||||||||||||
| Fractionated by hexane | 12a±0.21 | 16ab±0.24 | 22b±0.28 | 21b±0.28 | 33c±0.36 | 46d±0.43 | 28bc±0.31 | 36c±0.36 | 71e±0.69 | ||||||||||||||||||
| Fractionated by ethanol | 8a±0.03 | 18c±0.23 | 20c±0.23 | 11ab±0.19 | 29d±0.31 | 41e±0.43 | 25d±0.29 | 47e±0.43 | 67f±0.61 | ||||||||||||||||||
| Fractionated by water | 0a±0.000 | 0a±0.000 | 0a±0.000 | 0a±0.000 | 0a±0.000 | 0a±0.000 | 0a±0.000 | 0a±0.000 | 0a±0.000 |
Table 1: Effect of the raw extract of _Laminaria digitata_ and three soluble fractions by hexane, ethanol and water on mycelia gr
a Values with different letters for each row are significantly different according to DMRT at a probability P ≤ 0.05 level. Table 1: Effect of the raw extract of Laminaria digitata and three soluble fractions by hexane, ethanol and water on mycelia growth inhibition index (MGI%) of Rhizopus stolonifer. Activity of each extract applied at three increasing concentrations (from 10 to 30 g L-1) was tested by evaluation of MGI% after 1, 3 and 5 days of incubation at 18 ± 1 °C into Petri plates filled with potato dextrose agar media. Each value, ranging from 0% (no inhibition) to 100% (total inhibition), is the pooled mean ± SD of three replicates with 10 plates for each analyzed by two-way ANOVA.
| Concentration of extract (g L-1) | |||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10 | 20 | 30 | |||||||||||||||||||||||||||||||||
| Extract | |||||||||||||||||||||||||||||||||||
| Incubation time (hours) | |||||||||||||||||||||||||||||||||||
| 2 | 10 | 18 | 24 | 2 | 10 | 18 | 24 | 2 | 10 | 18 | 24 | ||||||||||||||||||||||||
| Raw | 15a±0. 21 | 21ab±0 .28 | 39c±0. 36 | 52d±0. 49 | 22ab±0 .28 | 34c±0. 36 | 57d±0. 51 | 75ef±0. 71 | 33c±0. 36 | 51d±0. 49 | 68e±0. 66 | 95g±0. 88 | |||||||||||||||||||||||
| Fraction ated by hexane | 10a±0. 21 | 17ab±0 .21 | 28c±0. 31 | 42d±0. 43 | 14a±0. 21 | 28c±0. 31 | 49d±0. 47 | 68ef±0. 61 | 23c±0. 27 | 45d±0. 41 | 61e±0. 61 | 82g±0. 75 | |||||||||||||||||||||||
| Fraction ated by ethanol | 7a±0.0 3 | 21c±0. 28 | 31d±0. 36 | 40e±0. 36 | 14b±0. 21 | 28cd±0 .31 | 42e±0. 37 | 57f±0. 51 | 13b±0. 21 | 38d±0. 39 | 51f±0. 46 | 69g±0. 61 | |||||||||||||||||||||||
| Fraction ated by water | 0a±0.0 00 | 1a±0.0 02 | 2a±0.0 03 | 1a±0.0 02 | 0a±0.0 00 | 0a±0.0 00 | 1a±0.0 02 | 2a±0.0 03 | 0a±0.0 00 | 0a±0.0 00 | 3a±0.0 04 | 2a±0.0 03 |
Table 2: Effect of the raw extract of _Laminaria digitata_ and three soluble fractions by hexane, ethanol and water on sporangia
a Values with different letters for each row are significantly different according to DMRT at a probability P ≤ 0.05 level. Table 2: Effect of the raw extract of Laminaria digitata and three soluble fractions by hexane, ethanol and water on sporangia germination suppression ratio (SG%) of Rhizopus stolonifer. Activity of each extract applied at three increasing concentrations (from 10 to 30 g L-1) was tested by assessment of SG% after 2, 10, 18 and 24 hours of incubation at 18 ± 1°C into micro-well plates filled with potato dextrose broth medium. Each value, ranging from 0% (no suppression) to 100% (total suppression), is the pooled mean ± SD of three replicates of 5 glass slides taken from each micro-well analyzed by two-way ANOVA.
Mycelia growth inhibition index a : 1) - Extract concentration (g L−1) 2 41.0 0.02 2) - Incubation time (day) 2 28.2 0.03 - Extract concentration × incubation time 4 3.8 0.02 Sporangia germination suppression ratio b : 3) - Extract concentration (g L−1) 2 48.6 0.01 4) - Incubation time (hour) 3 32.7 0.02 - Extract concentration × incubation time 6 4.7 0.03 Strawberry fruit soft rot inhibition index c : 5) - Extract concentration (g L−1) 2 35.6 <0.01 6) - Treatment type 1 7.6 <0.01 Effect df d F P-value De Corato U. Postharvest Control of Strawberry Fruit Soft Rot by Laminaria Digitata Extracts. J Agri Res 2019, 4(1): 000217.
| - Extract concentration × treatment type | 2 | 9.8 | <0.01 |
|---|
Table 3: Synthetic values (df and _F_) regarding to three different two-way ANOVA analysis for mycelia growth inhibition index, s
aExtract concentration (10, 20 and 30 g L−1) and incubation time (1, 3 and 5 days) are the two factors considered on mycelia growth inhibition index. bExtract concentration (10, 20 and 30 g L−1) and incubation time (2, 10, 18 and 24 hours) are the two factors considered on sporangia germination suppression ratio. c Extract concentration (10, 20 and 30 g L−1) and treatment type (preventive and curative) are the two factors considered on strawberry fruit rot suppression index. d Degree of freedom. Table 3: Synthetic values (df and F) regarding to three different two-way ANOVA analysis for mycelia growth inhibition index, sporangia germination suppression ratio, and fruit soft rot inhibition index at P-value ≤ 0.05 after application of Laminaria digitata raw extract performed by in vitro and in vivo experiments.
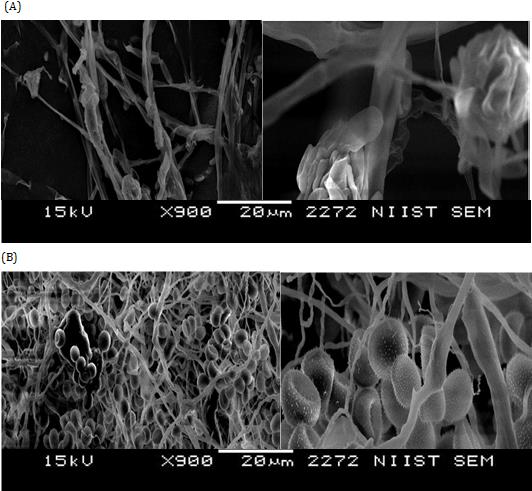
Scanning Electron Microscopy
Figure 1 shows the main morphological changes in R. stolonifer strain grown in PDB amended with 20 g L-1 L. digitata raw extract with respect to un-treated control after three days of incubation. The supplementation of extract: (i) reduced mycelia growth by retarding sporangiophore and sporangia formation, (ii) induced alteration in morphology of the sporangiophore which remained without sporangia and (iii) highly altered mycelia development with bare of sporangiospores and few collapsed sporangia (Figure 1A). Whereas, the control treated with the buffer alone showed normal sporangia and sporangiophore maturation, and hyphae growing (Figure 1B).
In vivo Effect
75%; but it resulted 3%, 16% and 23% in curative treatment by applying 10, 20 and 30 g L-1 of extract, respectively (Figure 2). The chemical treatment with Fenhexamid suppressed the strawberries soft rot of 90% in preventive treatment, whereas it was 72% in those curative (Figure 2). No suppressive effect applying the buffer without extract was found. Strawberry fruit soft rot inhibition index was significantly affect by the raw extract concentration applied over strawberries wound and treatment type (Table 3) since significant interactions between these two factors were shown (P-value < 0.05).
During the first day of soft rot infection, the strawberry wounds treated with the buffer alone in the control plot were covered with thin, fluffy and cotton-like fungal structures. Sporulation form represented by sporangiophore and sporangia appeared after inoculation for two days as a dark mass with black sporangia at their tips which covers the entire fruit wound. However, the typical symptoms of soft rot were observed after three days from artificial infection appearing as a deliquescence mass with loss of consistency and texture on strawberries tissue. The inhibition of fruit decay after preventive treatment with L. digitata raw extract was 28%, 47% and $$ \mathrm {d} = \mathrm {d} _ {1} + \mathrm {d} _ {2} + \mathrm {d} _ {3} + \dots + \mathrm {d} _ {n} $$
100 a
Figure 2: Soft rot suppression index on strawberries cv. Camarosa inoculated with Rhizopus stolonifer after three days of incubation at 18 ± 2 °C and 96 ± 2% RH in the dark after preventive and curative treatments with 10 g L−1 (TrL-10), 20 g L−1 (TrL-20), and 30 g L−1 (TrL-30) Laminaria digitata raw extract compared to 1.2 g L−1 Fenhexamid-50% (Con-Fen). Values ranging from 0% (no inhibition) to 100% (total inhibition) are the pooled mean of two experiments whenever each of them was carried out with six replicates per treatment and thirty strawberries for each replication with two wounds per fruit. Bar indicates the SD of the mean. For each treatment type, values with different letters are significantly different according to DMRT at a probability P ≤ 0.05 levels.
Peroxidase Activity
As regards to measurement of POD activity in strawberry fruit, a significant absorbance increasing in L. digitata-treated inoculated fruit was seen yet after one day of incubation (7.04 ΔOD420 g-1 min-1) when compared with that in untreated-inoculated strawberries (4.21 ΔOD420 g-1 min-1) (Table 4). While, the untreated-healthy fruit showed a significant lower change of POD activity (1.32 ΔOD420 g-1 min-1) than the untreated-inoculated TrL-10 TrL-20 $$ \mathrm {d} = \mathrm {d} _ {1} + \mathrm {d} _ {2} + \mathrm {d} _ {3} + \dots + \mathrm {d} _ {n} $$ TrL-30 Con-Fen a
| Preventive | c | 70 |
| Curative | c | 20 |
Table 5: Peroxidase (POD) activity in strawberries cv. Camarosa previously treated with 30 g L−1 _Laminaria_ _digitata_ raw extra
b c strawberry fruit after one day from pathogen inoculation (Table 4). It is also due to underline that differences in POD activity in L. digitata-treated inoculated fruit were more noticeable after three days of incubation (41.66 ΔOD420 g-1 min-1) than both controls, highlighting a significant increased POD activity at the end of incubation time.
| Experimental plot | 1 d | 3 d | ||||||
| L. digitata-treated inoculated strawberries | 7.04a ± 0.8a | 41.66a ± 1.5 | ||||||
| Untreated-inoculated strawberries | 4.21b ± 0.2 | 9.86b ± 0.8 | ||||||
| Untreated-healthy strawberries | 1.32c ± 0.09 | 2.2c ± 0.1 |
Table 6: Peroxidase (POD) activity in strawberries cv. Camarosa previously treated with 30 g L−1 _Laminaria_ _digitata_ raw extra
aChange in absorbance on a fresh weight basis (ΔOD420 g-1 min1). Table 4: Peroxidase (POD) activity in strawberries cv. Camarosa previously treated with 30 g L−1 Laminaria digitata raw extract and then inoculated with Rhizopus stolonifer. Assessment of POD activity was carried out after one day and three days (d) of incubation at 18 ± 2 °C and 98% RH in the dark. Values are the pooled mean ± SD of three replicates of 10 fruit for each analyzed by one- way ANOVA. For each column, values followed by different letters are significantly different according to DMRT at a probability P ≤ 0.05 level.
Discussion and Conclusions
In vitro experiments performed with the fractionated extracts of L. digitata by hexane and ethanol suggest that inhibition on mycelia growth and suppression on sporangia germination could be attributed to a direct toxicity of bioactive molecules, such as the fatty acids and phenolic compounds present in extracts purified by hexane and ethanol, respectively. Our findings support this hypothesis because both hexane- and ethanol-soluble extracts exerted an antifungal effect very similar either among them either to those seen testing the raw extract. This hypothesis is also supported by the chemical analyses carried out by Løvstad Holdt S, et al. which have detected higher content of fatty acids (lipids) and phenolic compounds (mainly phlorotannins) in raw extracts of L. digitata purified by hexane and ethanol, respectively [39]. In addition, efficacy of treatment significantly interacted with the extract concentration because suppression of mycelia growth and sporangia germination increase as the dose of extract added into growing media increased by a dose-dependent manner, at least in the concentration range considered here. Our findings showed that inhibition significantly interacted with the incubation time since it increases from the first to the last set point in a time-dependent manner, such showing that extracts remain really efficacy in inhibiting fungal growth during all incubation time. Similar conclusions were obtained by in vivo experiments performed with the raw extract during preventive treatment, whenever the application of 30 g L−1 extract shows the strongest efficacy in suppressing fruit decay with respect to curative treatment. Therefore, the efficacy of the treatments was related to dose of extract applied whenever an evident ‘dose-effect’ was observed. Findings coming from these experiments also highlight a very competitive efficacy of the raw extract when compared to Fenhexamid in preventive treatment, whenever soft rot development in L. digitata-treated strawberries after three days of incubation was comparable to that seen in Fenhexamid-treated strawberries after five days (data not shown) confirming persistency of antifungal efficacy of the raw extract until five days from pathogen inoculation. However, it is due to underline that postharvest application of Fenhexamid is banned by the Italian and European laws (EC Regulation No. 1107/2009, repealing Directive 91/414/EEC), thus it has been used in this work as a reference for experimental purpose only. In addition, our findings showed that strawberries soft rot suppression significantly interacted either with the extract dose or treatment type, since a remarkable increasing of suppression was seen as the concentration of extract applied over the wounds increased, as well as through gone from the curative treatment into preventive. Although seaweed extract contains a relevant number of bioactive compounds with antimicrobial, antiviral, anticancer, antioxidant, and antifungal properties that have been studied for several applications, nonetheless the direct effect of them on strawberry fungal pathogens, or their possible resistance induction effect on strawberry plants, has not been thoroughly investigated in depth [22, 39]. On the whole, findings coming from this work show that a direct antifungal activity exerted by the raw extract of L. digitata could be attributed to its content of fatty acids according to De Corato U, et al., and that preventive treatments are really competitive to control postharvest soft rot of strawberries in comparison to Fenhexamid [32]. Authors have investigated on the postharvest antifungal property of crude extracts obtained by the supercritical carbon dioxide technique from a collection of five seaweed species, included L. digitata, in suppressing B. cinerea, Monilinia laxa (Aderh. & Ruhland) Honey, and Penicillium digitatum (Pers.) Sacc. on strawberries, peaches and lemons, respectively [32]. Findings came from the present work are nevertheless divergent from those given by De Corato U, et al. in relation to efficacy of the ethanolic fraction from L. digitata, resulting significantly less suppressive to B. cinerea and M. laxa than hexanic fraction [32]. This discordance is probably due to the different extractive techniques used, being employed the supercritical carbon dioxide technique to maximize extraction of lipid (fatty acids) rather than phenolic substance (phlorotannins) at the fixed conditions of pressure and temperature when compared to methods that uses organic solvents. In fact, the extraction technique used here is less selective and more suitable to extract a wider range of biologically active substances without preferences among them whenever mixtures of polar and un-polar solvents were employed as extractives at the proper rates [33]. Thus, the phenolic fraction purified by ethanol could also be considered a reliable candidate to suppress mycelia growth and sporangia germination of R. stolonifer acting in synergy with the fatty acids. To explore the most probable mechanisms of action by which fatty acids and phenolic molecules can inhibit mycelia growth and sporangia germination, we have seen that the morphology of sporangia and hyphae of R. stolonifer exposed to L. digitata raw extract shows remarkable alterations as reduction of sporangia number, sporangia swelling, sporangia wall collapse, hyphae blowing, and more alterations of their walls. In addition, our findings result very similar to those reported by Salem EA, et al. in a R. stolonifer pathogenic strain grown in PDB amended with 1.5 g L-1 calcium chloride, or 2 mL L-
1 lemongrass essential oil, or a combination of them [20]. Damaging mechanisms induced by longer exposure to essential oil vapour have also been reported in literature, such as a partition of lipid layer of the cell membrane due to their hydrophobic nature and affection of permeability of the cell membrane that cause leakage of cell components [40, 41, 42, 43]. In addition, several ultra-structural damages on the fungal cells caused to their walls have also been reported in literature as a result of the direct interaction of many bioactive metabolites from plant and macroalgae, including fatty acids and phenolic substances, with the enzymes responsible to cell wall synthesis [44]. On the other hand, in vitro inhibition on mycelia growth and sporangia germination were not detected by applying the water-soluble extract fraction, such suggesting that the aqueous extract containing mainly polysaccharides could be not directly involved in a direct inhibition against mycelia development and sporangia germination. But, at the same time, a significant increasing of POD activity in L. digitata-treated inoculated strawberries was earlier found from the artificial inoculation, and overall till the end of incubation time whenever these differences were more noticeable if compared to untreated-inoculated fruit. Moreover, healthy strawberries treated with the sterile buffer alone showed a significant POD decreasing with respect to untreated-inoculated fruit. These significant increments of POD activity yet after one day from treatment, as well as an increased POD activity after three days of incubation, could be both related to activation of systemic acquired resistance (SAR) around tissue of the inoculated strawberries. This hypothesis is supported by the fact that artificial inoculation with the pathogen without extract induced a low absorbance change with respect to L. digitata-treated inoculated strawberries, while healthy fruit showed the lowest absorbance change in comparison to others plots. In fact, might be reasonable affirm that polysaccharides and oligosaccharides (as laminarans, fucoidans, and alginates) into L. digitata raw extract could elicit host defense responses or signal transduction molecules in planta through an increased hydrogen peroxide production by acting as oxidant agents rather than as chemical toxics to pathogen into fruit tissue [39, 45, 46, 47]. Particularly laminarin, a storage polysaccharide (β-1,3-glucan) isolated for the first time from the cell walls of L. digitata, is a known elicitor which promotes defense mechanisms responses in planta including Vitis vinifera L. against B. cinerea and Plasmopara viticola (Berk. & M.A. Curtis) Berl. & De Toni [48, 49, 50, 51]. In addition, a commercial formulate containing 45 g L-1 purified laminarin ( ‘Vacciplant’) is currently used in organic farming systems for controlling postharvest gray mold by B. cinerea and soft rot by Rhizopus spp. on strawberries [52, 53]. More recently, authors have investigated on the reliable application of laminarin associated to calcium oxide to control grape powdery mildew on Moscato grape, such demonstrating that the association of laminarin and calcium oxide shows a significant reduction of powdery mildew symptoms by comparing them to efficacy of sulphur [54]. Nevertheless, is also due to underline that other defense-related enzymes such as chitinase, β-1,3-glucanase, polyphenol oxidase, phenylalanine ammonia lyase, and lipoxygenase might be involved in suppression and that should be considered in depth in future researches of this topic. In conclusion, in vitro antifungal activity of L. digitata raw extract and purified by hexane, ethanol and water, as well as in vivo investigations on antifungal efficacy of L. digitata raw extract to postharvest Rhizopus soft rot of strawberry fruit, were both evaluated. Findings of this work have gave promising results in relation to potential use of L. digitata raw extract against R. stolonifer attacking strawberry fruit as alternative to Fenhexamid during preventive treatment. Nevertheless, fewer number of referred papers on antifungal activity of L. digitata raw extract to postharvest Rhizopus soft rot of strawberry fruit have been found in literature, which thus have not allowed an adequate amount of comparisons with the existing references. However, as main conclusion, we can affirm that a new alternative mean for effective control postharvest soft rot of strawberries is needed to obtain marketable commodities without fungicide residues. L. digitata raw extract could be used in preventive treatment as a suitable control mean due to its higher competitiveness than the banned synthetic fungicides. A direct antifungal activity to sporangia germination and hyphae development elicited by both raw and fractionated extracts by hexane and ethanol could be attributed to toxicity due to fatty acids and phenolic compounds extracted by mixtures of polar and un-polar solvents at the appropriate rates. Nevertheless, an increased POD activity probably elicited by the water- soluble polysaccharides should be related to activation of SAR in suppressing postharvest soft rot on strawberries. Although further large scale trials are required, nonetheless one possible way to reach this goal could be the use of L. digitata extract produced into a marine biorefineries under industrial scale.
References
-
Khoshnevisan B, Rafiee S, Mousazadeh H (2013) Environmental impact assessment of open field and greenhouse strawberry production. Eur J Agron 50: 29-37.
-
Khoshnevisan B, Rafiee S, Mousazadeh H (2014) Application of multilayer adaptive neuro-fuzzy inference system for estimation of greenhouse strawberry yield. Measurement 47: 903-910.
-
De Cicco V, Bertolini P, Salerno MG (2008) Patologia post-raccolta dei prodotti vegetali. Piccin Press, Bologna, Italy, pp: 188.
-
Husaini AM, Neri D (2016) Strawberry growth, development and diseases. CABI, Wallingford.
-
Feliziani E, Romanazzi G (2016) Postharvest decay of strawberry fruit: Etiology, epidemiology, and disease management. J Berry Research 6: 47-63.
-
Aliasgarian S, Ghassemzadeh HR, Moghaddam M, Ghaffari H (2013) Mechanical damage of strawberry during harvest and postharvest operations. J World Appl Sci 22(7): 969-974.
-
Maas JL (1998) Compendium of strawberry diseases. Second Edition, APS Press, St. Paul, MN, USA, pp: 138.
-
Boer SHJ, Boyd-Wilson KSH, Petley M, Walter M (2009) Influence of cold-storage temperatures on strawberry leak caused by _Rhizopus_ spp. N Z Plant Protect 62: 243-249.
-
Bautista-Baños S, Bosquez-Molina E, Barrera-Necha LL (2014) _Rhizopus stolonifer_ (Soft rot). _In:_ Bautista- Baños S (Ed.), Postharvest decay of fruit and vegetables: Control strategies. Elsevier Press, pp: 383.
-
Ippolito A, Nigro F, Romanazzi G, Campanella V (1997) Field application of _Aureobasidium pullulans_ against _Botrytis_ storage rot of strawberry. _In:_ Bertolini P, Sijmons PC, Guerzoni ME, Serra F (Eds.), No conventional methods for the control of post- harvest disease and microbiological spoilage. Proceedings of the COST 914-COST 915, Bologna, Italy, pp: 127-133.
-
Droby S, Wisniewski M, Macarisin D, Wilson C (2009) Twenty years of postharvest biocontrol research: is it time for a new paradigm? Postharvest Biol Technol 52(2): 137-145.
-
Sanzani SM, Nigro F, Mari M, Ippolito A (2009) Innovations in the control of postharvest diseases of fresh fruits and vegetables. Arabian J Plant Prot 27(2): 240-244.
-
Feliziani E, Romanazzi G (2018) Preservation of berries. Encyclopedia of Food Security and Sustainability.
-
Yang X, Jiang X. (2015) Antifungal activity and mechanism of tea polyphenols against _Rhizopus_ _stolonifer_. Biotechnol Lett 37(7): 1463-1472.
-
Mari M, Bautista-Baños S, Sivakumar D (2016) Decay control in the postharvest system: Role of microbial and plant volatile organic compounds. Postharvest Biol Technol 122: 70-81.
-
Gatto MA, Ippolito A, Linsalata V, Cascarano NA, Nigro F, et al. (2011) Activity of extracts from wild edible herbs against postharvest fungal diseases of fruit and vegetables. Postharvest Biol Technol 61(1): 72-82.
-
Gatto MA, Sanzani SM, Tardia P, Linsalata V, Pieralice M, et al. (2013) Antifungal activity of total and fractionated phenolic extracts from two wild edible herbs. Nat Sci 5(8): 895-902.
-
De Corato U, Maccioni O, Trupo M, Di Sanzo G (2010) Use of essential oil of _Lauris nobilis_ obtained by means of a superficial carbon dioxide technique against postharvest spoilage fungi. Crop Prot 29(2): 142-147.
-
Hadian J, Ghasemnezhad M, Ranjbar H, Frazane M, Ghorbanpour M (2008) Antifungal potency of some essential oils in control of postharvest decay of strawberry caused by _Botrytis cinerea_, _Rhizopus_ _stolonifer_ and _Aspergillus niger_. J Essent Oil Bear Pl 11(5): 553-562.
-
Salem EA, Youssef K, Sanzani SM (2016) Evaluation of alternative means to control postharvest _Rhizopus_ rot of peaches. Sci Hort 198: 86-90.
-
Tuhy L, Chowanska J, Chojnacka K (2013) Seaweed extracts as biostimulants of plant growth: A review. Ckemik 67(7): 636-641.
-
Righini I, Roberti R, Baraldi E (2018) Use of algae in strawberry management. J Appl Phycol 30(6): 3551- 3564.
-
Calvo P, Nelson L, Kloepper JW (2014) Agricultural uses of plant biostimulants. Plant Soil 383(1-2): 3-41.
-
Craigie JS (2011) Seaweed extract stimuli in plant science and agriculture. J Appl Phycol 23(3): 371-393.
-
Ibrahim WM, Refaat MA, Khaulood AH, Makram AS (2014) Role of _Ulva lactuca_ extract in alleviation of salinity stress on wheat seedlings. Sci World J 2014: 1-11.
-
Latique S, Elouaer MA, Chernane H, Hannachi C, Elkaou M (2014) Effect of seaweed liquid extract of _Sargassum vulgare_ on growth of durum wheat seedlings (_Triticum durum_ L) under salt stress. Int J Innovat Appl Studi 7(4): 1430-1435.
-
Jayaraman J, Norrie J, Punja ZK (2011) Commercial extract from the brown seaweed _Ascophyllum_ _nodosum_ reduces fungal diseases in greenhouse cucumber. J Appl Phycol 23(3): 353-361.
-
Hernández-Herrera RM, Virgen-Calleros G, Ruiz- López M, Zañudo-Hernández J, Délano-Frier JP, et al. (2014) Extracts from green and brown seaweeds protect tomato (_Solanum lycopersicum_) against the necrotrophic fungus _Alternaria solani_. J Appl Phycol 26(3): 1607-1614.
-
Roberti R, Righini H, Pérez Reyes C (2016) Activity of seaweed and cyanobacteria water extracts against _Podosphaera xanthii_ on zucchini. Ital J Mycol 45: 66- 77.
-
Esserti S, Smaili A, Rifai LA, Koussa T, Makroum K, et al. (2017) Protective effect of three brown seaweed extracts against fungal and bacterial diseases of tomato. J Appl Phycol 29(2): 1081-1093.
-
Washington WS, Engleitner S, Boontjes G, Shanmuganathan N (1999) Effects of fungicides, seaweed extract, tea tree oil and fungal agents on fruit rot and yield in strawberry. Australian J Exp Agric 39: 487-494.
-
De Corato U, Salimbeni R, De Pretis A, Avella N, Patruno G (2017) Antifungal activity of crude extracts from brown and red seaweeds by a supercritical carbon dioxide technique against fruit postharvest fungal diseases. Postharvest Biol Technol 131: 16-30.
-
Kulandaivel S (2014) Study on phytochemical and antimicrobial activity of some selected marine seaweeds against human and fish pathogens. Innovare J Ayurvedic Sci 2(1): 1-5.
-
Khanzada AK, Shaikh W, Kazi TG, Kabir S, Soofia S (2007) Antifungal activity, elemental analysis and determination of total protein of seaweed _Solieria_ _robusta_ (greville) kylin from the coast of Karachi. Pakistan J Bot 39(3): 931-937.
-
Lima JN, Silva GB, Cordeiro CS, Lins MM, Oliveira L, et al. (2014) Isolation, identification and phenotypiccharacterisation of _Rhizopus_ strains isolated from Caatinga soil in Pernambuco, Brazil. _In:_ Industrial, Medical and Environmental Applications of Microorganisms: Current Status and Trends. Proceedings of the V International Conference on Environmental, Industrial and Applied Microbiology (BioMicroWorld 2013).Wageningen (Holland), Academic Publishers, pp: 119.
-
Shahi SK, Shukla AC, Bajaj AK, Midgely G, Dikshit A (1999) Broad spectrum antimycotic drug for the control of fungal infection in human beings. Curr Sci 76(6): 836-839.
-
Harley MM, Fergusen IK (1990) The role of the SEM in pollen morphology and plant systematics. _In:_ Claugher D (Ed.), Scanning Electron Microscope in Taxonomy and Functional Morphology. Systematics Association Special, vol.41. Clarendon Press, Oxford, UK, pp: 45-68.
-
Hammerschmidt R, Nuckles EM, Kuc J (1982) Association of enhanced peroxidase activity with induced systemic resistance of cucumber to _Colletotrichum lagenarium_. Physiol Plant Pathol 20(1): 77-82.
-
Holdt SL, Kraan S (2011) Bioactive compounds in seaweed: functional food applications and legislation. J Appl Phycol 23(3): 543-597.
-
Soylu EM, Soylu S, Kurt S (2006) Antimicrobial activities of the essential oils of various plants against late blight disease agent _Phytophthora infestans_. Mycopathologia 161(2): 119-128.
-
Laird K, Phillips C (2011) Vapour phase: a potential future use for essential oils as antimicrobials? Lett Appl Microbiol 54(3): 169-174.
-
da Cruz CL, Pinto VF, Patriarca A (2013) Application of plant derived compounds to control fungal spoilage and mycotoxin production in foods. Int Food Microbiol 166(1): 1-14.
-
Shao X, Wang H, Xu F, Cheng S (2013) Effects and possible mechanisms of tea tree oil vapor treatments on the main disease in postharvest strawberry fruit. Postharvest Biol Technol 77: 94-101.
-
Rasooli I, Rezaei MB, Allameh A (2006) Growth inhibition and morphological alterations of _Aspergillus niger_ by essential oils from _Thymus_ _eriocalyx_ and _Thymus x-porlock_. Food Control 17(5): 359-364.
-
Walters D, Walsh D, Newton A, Lyon G (2005) Induced resistance for plant disease control: maximizing the efficacy of resistance elicitors. Phytopathology 95(12): 1368-1373.
-
Imlay JA, Linn S (1988) DNA damage and oxygen radical toxicity. Science 240(4857): 1302-1309.
-
Tatsuzawa H, Maruyama T, Misawa N, Fujimori K, Hori K, et al. (1998) Inactivation of bacterial respiratory chain enzymes by singlet oxygen. FEBS Letters 439(3): 329-333.
-
Labarre A, Orieux R (2010) Plant defences stimulation: Laminarin, a natural compound for the plant protection. Julius-Kühn-Arch 428: 117.
-
Bernardon MA, Joubert JM (2012) La laminarine contre la tavelure (_Venturia inaequalis_) du pommier e le _Gleosporium_ des pommes (_Gleosporium album_ et _perenans_). Afpp-10e Conférence Internationale Sur Les Maladies Des Plantes Tours (France), pp: 630- 639.
-
Aziz A, Poinssot B, Daire X, Adrian M, Bazier A, et al. (2003) Laminarin elicits defense responses in grapevine and induces protection against _Botrytis_ _cinerea_ and _Plasmopara viticola_. Mol Plant Microbe Interact 16(12): 1118-1128.
-
Trouvelot S, Varnier AL, Allègre M, Mercier L, Baillieul F, et al. (2008) A beta-1,3 glucan sulfate induces resistance in grapevine against _Plasmopara viticola_ through priming of defense responses, including HR- like cell death. Mol Plant Microbe Interact 21(12): 232-243.
-
De Miccolis R, Angelini M, Pollastro S, Faretra F (2009) Induction of plant resistance in crop protection against fungal diseases. _In:_ Petria, Resistenza indotta per il controllo di malattie delle piante 19(1): 57-59.
-
Santini M, Landi L, Feliziani E, Murolo S, Romanazzi G (2009) Use of resistance inducers for the control of postharvest gray mold and _Rhizopus_ rot in strawberry. _In:_ Petria, Resistenza indotta per il controllo di malattie delle piante 19(1): 73-76.
-
Pugliese M, Monchiero M, Gullino ML, Garibaldi A (2018) Application of laminarin and calcium oxide for the control of grape powdery mildew on _Vitis vinifera_ cv. Moscato. J Pl Dis Prot 125(5): 477-482.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note