Morphological and Molecular Detection of Spongospora subterranea on Plant Harvest in Infected Powder Scab Plots in Antioquia
Potato powdery scab is a disease that negatively affects both roots, causing gall formation, and tubers, resulting in brown pustules. Additionally, it leads to the development of resistance structures that can persist in the soil for several years. Globally, it is one of the most limiting diseases for potato cultivation. This study aimed to identify the species present in areas with a historical occurrence of powdery scab and to evaluate their role as alternative hosts for the pathogen. Samples were collected in the municipalities of Santa Rosa de Osos and La Unión (Ant- Col). The presence of structures (plasmodia and cystosori) associated with the pathogen in the roots was determined using trypan blue staining followed by optical microscopy analysis. Morphological identification revealed twenty-six families with pathogen structures, including Acmella mutisii, Amaranthus dubius, Bidens pilosa, Galinsoga quadriradiata, Holcus lanatus, Hypochaeris radicata, Juncus microcephalus, Oxalis corniculata, Pennisetum clandestinum, Phytolacca bogotensis, Poa annua, Polygonum nepalense, Raphanus raphanistrum, Rumex crispus, Sonchus oleraceus, Trifolium repens, Verbena litoralis, along with Fragaria x ananassa, Solanum tuberosum, and Zea mays. Molecular analysis using RT-PCR for the detection of Plasmodiophoromycetes revealed the presence of Spongospora subterranea in Solanum tuberosum; Polymyxa betae in S. tuberosum, O. corniculata, and T. repens; and Polymyxa graminis in Z. mays. This comprehensive study highlights the importance of crop rotation and weed control to disrupt the pathogen’s lifecycle as effective strategies to mitigate the impact of the disease. It emphasizes the role of alternative hosts and their substantial contribution to elevated inoculum levels.
Introduction
Potato powdery scab is caused by Spongospora subterranea f. sp. subterranea, a protozoan belonging to the Plasmodiophorales order. This order constitutes a monophyletic group, and its members share common characteristics, including cruciform nuclear division, zoospores with two anterior flagella, an ameboidal phase, multinucleated protoplast, plasmodia, obligatory intracellular parasitism, and the formation of resistance spores within cysts. Currently, S. subterranea cannot be cultured in vitro [1, 2].
In Colombia, the first report of potato powdery scab dates back to 1965 [3]. Presently, it has spread to regions such as Boyacá, Cundinamarca, Nariño, and Antioquia, impacting tuber production by 50 to 80%, depending on the crop’s age at the time of infection. This situation results in substantial economic losses for the agro-industrial sector, posing phytosanitary challenges due to the pathogen’s dissemination to disease-free areas, and hindering certification for seed producers [4, 5].
This disease is prevalent in nearly all potato-producing regions worldwide, resulting in significant losses in both seed and consumption tubers [6]. On tubers, it manifests as open pustules ranging from light to dark brown, typically circular in shape, thereby diminishing tuber quality [7, 8]. In the roots, it gives rise to distorted galls and nodules in the stolon. Initially, the galls appear whitish, but as they reach physiological maturity, they darken due to the brown color of the resistance structures’ walls [9]. Notably, the sporeballs can persist in the soil for extended periods [10].
According to Balendres MA, et al. [5] sporeballs can germinate in the presence of host plants or environmental stimuli, initiating the disease cycle early on, irrespective of their maturity stage.
Asserts that a high level of root infection negatively impacts water and nutrient intake, leading to reduced yield production [5, 11]. In advanced disease states, the roots disintegrate, releasing cystosori massively, becoming a long- lasting soil inoculum source, thereby making it a challenging disease to control. Additionally, S. subterranea serves as the vector for the “Potato Mop Top Virus” (PMTV), residing in the pathogen’s spores for several years [12, 13]. Currently, there is no chemical treatment available for disease control. Tsror (Lahkim) L, et al. [14] discovered that the use of sodium metanum and chloropicrin proved highly effective in controlling the disease under field conditions compared to the control treatment. However, at the end of the growth cycle, disease levels increased again without significant differences between all treatments.
The range of hosts reported for S. subterranea is extensive, encompassing species from families such as Aizoaceae, Apiaceae, Asteraceae, Boraginaceae, Brassicaceae, Caryophyllaceae, Chenopodiaceae, Coniferae, Fabaceae, Lamiaceae, Malvaceae, Papaveraceae, Plantaginaceae, Poaceae, Polygonaceae, Ranunculaceae, Resedaceae, Solanaceae, Urticaceae, and Zygophyllaceae [12, 14, 15, 16].
Evaluated common weed plant species in Danish potato fields cultivated in hydroponic systems infested with zoospores of S. subterranea. Through DAS-ELISA and visible symptom presence, they found that 13 species were infected with zoosporangia of S. subterranea, although without galls in the roots [17].
Determined the host range of S. subterranea in different crops and some weed plants in the northeastern United States. The study evaluated 26 species among 10 families of monocotyledonous and dicotyledonous plants, discovering 16 species susceptible to S. subterranea, with 12 new hosts reported. Root galls were identified in six species, and cystosori were found in three species. These results suggest that S. subterranea has a broad host range in its zoosporangial phase, but only in a few hosts does the pathogen complete its life cycle, producing cystospores [18].
The advancement of effective strategies for managing S. subterranea has been hindered by the absence of a rapid and efficient method for detecting pathogens in the soil [1]. Various alternatives for pathogen detection have been explored, ranging from the utilization of plants in infected soils for 4-5 weeks [15]. Microscopic observations [15, 19, 20]sequenced, and assessed for genetic variation. Two genetically distinct groups (I and II, Elisa tests [21]. The application of molecular PCR techniques with specific high- sensitivity primers [15, 22, 23], and real-time PCR. These tests can be conducted on both plant tissues and soil, enabling the early identification of the causative agent of powdery scab in soils and plants [24].
Monoclonal antibodies, as utilized by Merz U, et al. [21], demonstrated the capability to detect less than one cystosorus of S. subterranea, recognizing it in samples from various countries. This technique exhibited no cross- reactions with other plasmodiophoromycetes such as Plasmodiophora brassicae, Polymyxa graminis, Polymyxa betae, and different species of Streptomyces. Notably, this method allowed the discrimination of different infection levels in a soil sample. The objective of this study was to identify structures associated with S. subterranea in different weed and crop plant species using light microscopy techniques and PCR tests. The investigation focused on plants growing in four types of plots with a history of powdery scab in two municipalities of Antioquia.
Materials and Methods
Field samples were collected from the municipalities of Santa Rosa de Osos (El Roble, Aragón, and El Tres) and La Unión (Chuscalito, Quebrada Negra, and San Francisco) in Antioquia, Colombia. Four types of plots with a history of powdery scab were chosen for sampling: those planted with potatoes, freshly harvested plots, resting fields, and plots growing a crop different from potatoes. A maximum of 10 plants per species was sampled in each plot. The collected plant specimens were brought to the Gabriel Gutiérrez Villegas Herbarium at Universidad Nacional de Colombia “MEDEL” and the Joaquín Antonio Uribe “JAUM” Herbarium at the Medellin Botanical Garden for botanical identification (Figure 1).
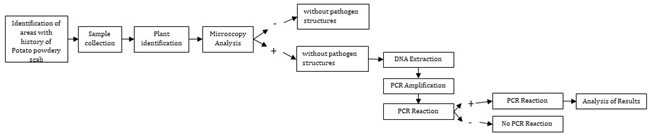
Microscopy Analysis
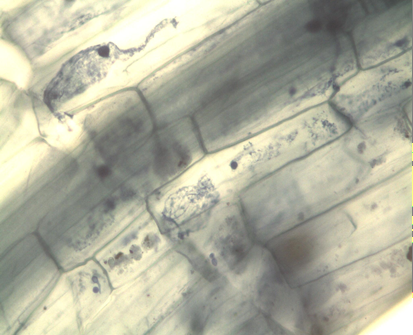
The collected samples underwent washing and immersion in a 10% KOH solution until laboratory analysis. To detect intracellular structures associated with S. subterranea, half of the roots were stained using the protocol with 0.05% trypan blue, modifying Irmayanti S, et al. [25] root clarification times as needed. The remaining half of the roots, designated for molecular analysis, were stored in bulk per species and dried at room temperature. For microscopic evaluation, five roots from each sample were placed on slides and observed at 10x and 40x magnifications under a microscope.
DNA Extraction
Following the observation of structures associated with S. subterranea or other plasmodiophoromycetes under microscopy, DNA extraction was performed for species where such structures were identified. Dry roots stored for each species sample were macerated into a fine powder using a mortar. DNA extraction was carried out using a commercial DNeasy Plant Mini Qiagen kit, following the manufacturer’s protocol. The obtained DNA was stored at -20°C for further analysis.
PCR Amplification
For the detection of S. subterranea f sp. subterranea, specific primers Spo8 - Spo9 [22], and SsF – SsR Qu X, et al. [26] were employed. These primers amplify regions of the DNAr, producing fragments of 390 and 434 bp, respectively. This approach allows differentiation between S. subterranea f. sp and S. nasturtii f. sp, as well as other Plasmodiophoromycetes like P. brassicae and P. graminis. Additionally, the primer pair 5.8S:32-for and 25-rev served as an internal amplification control, targeting a region between the 5.8S subunit and position 45 of the 28S subunit of ribosomal DNA in plants [27].
Simultaneously, to ensure that the observed structures were not caused by other microorganisms, the detection of Polymyxa graminis, Polymyxa betae, Plasmodiophora brassicae, S_. subterranea_ f.sp. nasturtii, and Sorosphaera veronicae was conducted. This was achieved using primers PNS1 - GRA2, PNS1 - BET1, NS1b – CR2, NS1b – WC1, and PNS1 – SV1, respectively (22)Peru, and Australasia. No sequence variation was detected between any of the Australasian or European collections with the exception of one from Inverness (Scotland. Furthermore, the ITS1 primer W White T, et al. [28] was employed along with CR2, WC1, GRA2, BET1, and SV1 to detect P. graminis, P. betae, P. brassicae, S. subterranea fsp. nasturtii, and S. veronicae, respectively. The primer sequences used in this study are detailed in Table 1.
| PRIMERS | SEQUENCE | SPECIES |
|---|---|---|
| 5.8s:32f - 25Rev (27) | GCATCGATGAAGAACGTAGC TATGCTTAAAYTCAGCGGGT | Plant (control) |
| SsR – SsF (26) | GCACGCCAATGGTTAGAGACG GTCGGTTCTACCGGCAGACC | S. subterranea. fsp. Subterranea |
| SPO8 - SPO9 (22) | CTGGGTGCGATTGTCTGTTG CACGCCAATGGTTAGAGACG | S. subterranea. fsp. Subterranea |
| *NS1b - CR2 | GGTTGATCCTGCCAGTAGTG TATGCCGCAGCAAAGCTC | Plasmodiophora brassicae |
| *NS1b - WC1 | GGTTGATCCTGCCAGTAGTG GCAGACAGATTTGACTCTGG | S. s. f.sp nasturtii |
| *PNS1 - GRA2 | GTTATCTGGTTGATCCTGCC GTTCCAAGAACCCGATGGAC | Polymixa gramminis |
| *PNS1 - BET1 | GTTATCTGGTTGATCCTGCC CGAATCGACTCTCATTGTCC | P. betae (Keskin) |
| *PNS1 - SV1 | GTTATCTGGTTGATCCTGCC GCCGAGAATCACATTCAACC | Sorosphaera veronicae |
| **ITS1 - CR2 | TCCGTAGGTGAACCTTGCGG TATGCCGCAGCAAAGCTC | P. brassicae |
| **ITS1 - WC1 | TCCGTAGGTGAACCTTGCGG GCAGACAGATTTGACTCTGG | S. s. f.sp nasturtii |
| **ITS1 - GRA2 | TCCGTAGGTGAACCTTGCGG GTTCCAAGAACCCGATGGAC | P. gramminis |
| **ITS1 - BET1 | TCCGTAGGTGAACCTTGCGG CGAATCGACTCTCATTGTCC | P. betae |
| **ITS1 - SV1 | TCCGTAGGTGAACCTTGCGG GCCGAGAATCACATTCAACC | S. veronicae |
| * (22) ** (28) |
Table 1: Primers and Sequences Used for the Molecular Detection of Different Plasmodioforomycetes, in Different Species that Pres
PCR Reaction
PCR reactions for primers 25Rev - 8s:32f; SSR – SSF; SPO8 - SPO9; NS1b - CR2; NS1b - WC1; PNS1 - GRA2; PNS1 - BET1; PNS1 - SV1; ITS1 - CR2; ITS1 - WC1; ITS1 - GRA2; ITS1 - BET1; and ITS1 - SV1 comprised 0.5 µL of each primer, 1 U of Taq DNA recombinant polymerase (Fermentas, Vilnius, Lithuania), 0.2 mM of each dNTP, 1X enzyme buffer, 1.8 mM MgCl2, 1 µL of DNA as the template, and ultra-pure sterile water for a total volume of 25 μL.
Amplifications were conducted using a thermocycler (Multigene, Labnet). The procedure included an initial denaturation at 98°C for 3 min, followed by 40 cycles of 94°C for 30 sec, 54°C for 30 sec, 72°C for 2:30 min, and a final extension period at 72°C for 10 min for primers 25Rev- 8s:32f; SSR – SSF; and SPO8 - SPO9 [22]. For the amplification of primers NS1b - CR2; NS1b - WC1; PNS1 - GRA2; PNS1 - BET1; and PNS1 - SV1; ITS1 - CR2; ITS1-WC1; ITS1 - GRA2; ITS1 - BET1; and ITS1 - SV1, the protocol included an initial denaturation at 98°C for 3 min, followed by 40 cycles of 94°C for 30 sec, 56°C for 30 sec, 72°C for 3 min, and a final extension period at 72°C for 10 min [22].
Electrophoresis after amplification, 5 µL of the reaction products were taken and separated by 1.5% agarose gel electrophoresis for 45 min at 70 V. EZ-visionTM Three (AMRESCO) was used as the colorant. The fragments were visualized under ultraviolet light using a Benchtop 3UVTM Trans illuminator and the digital analysis system BioDoc (IT TM Imaging System). Electrophoresis included a molecular weight marker of 100 bp (GeneRuler. DNA Ladder, ready-to- use, Fermentas) for comparison.
Analysis of Results
The evaluation of results occurred in two phases. Firstly, the observation focused on structures associated with the pathogen, including cystosori and plasmodia inside the roots, through microscopy. Samples displaying such structures underwent further assessment through molecular tests. In these tests, the presence of amplicons was visualized by PCR products in agarose gel, confirming the existence of specific plasmodiophoromycetes.
Results and Discussion
A total of 1087 samples were collected across the two municipalities, with 648 samples from La Unión, representing 46 plant species, and 439 from Santa Rosa de Osos, representing 24 species. Following microscopic analysis for the identification of structures associated with S. subterranea, such as plasmodia and cystosori within the roots, it was observed that 26 species showed structures associated with the pathogen (Table 2). In the municipality of La Unión, 88 out of 648 samples exhibited these structures, while in Santa Rosa de Osos, 33 out of 439 samples displayed structures linked to the pathogen (Tables 3 & 4). Seventeen species in La Unión and seven in Santa Rosa showed structures associated with the pathogen, resulting in a total of 26 species with the presence of pathogen structures between the two municipalities. Notably, symptoms of galls roots were not observed in any of the evaluated species. Structures found inside the roots of some of these species (Figure 2).
| Species | Common Name | Municipality |
|---|---|---|
| Asteraceae (undetermined species) | - | La Unión – Santa Rosa |
| Caryophyllaceae (undetermined species) | - | La Unión |
| Conyza bonariensis | Asthmaweed, Hairy fleabane. | La Unión |
| Cucurbita maxima | Pumpkin, Squash | La Unión |
| Cyperus ferax | Rusty flat-sedge, slender nutsedge | Santa Rosa |
| Eragrostis soratensis | Elastic grass, Wiry lovegrass | Santa Rosa |
| Gnaphalium pisatum | Winged cudweed | Santa Rosa |
| Holcus sp. | - | La Unión |
| Lepidium bipinnatifidium | Virginia pepperweed, poorman’s pepper | La Unión |
| Lepidium sp. | - | La Unión |
| Lepidium costaricense | Pepperweed, Peppercress | Santa Rosa |
| Malva silvestris | Little mallow, Cheeseweed | La Unión |
| Oxalis latifolia | Broadleaf woodsorrel, | La Unión |
| Phaseolus vulgaris | Snap bean, Garden bean | La Unión |
| Plantago major | Common plantain | La Unión |
| Poa sp. | - | La Unión |
| Rumex acetocella | Sheep, Sorrel | La Unión |
| Rumex sp. | - | La Unión |
| Senecio vulgaris | Common groundsel | La Unión |
| Sonchus asper | Prickly, Sowthistle | La Unión – Santa Rosa |
| Spergula arvensis | Abrejillo - Miona | La Unión – Santa Rosa |
| Spermacoce assurgens | Shrubby false buttonweed | Santa Rosa |
| Stellaria sp. | - | Santa Rosa |
| Sterilla sp. | - | La Unión |
| Taraxacum officinale | Blowballs, Common dandelion | Santa Rosa |
| Veronica javanica | - | La Unión |
Table 2: Species that did not Present Structures Associated with S. Subterranea in the Municipalities of Santa Rosa De Osos and L
| Family | Species | Lot Type | |||
|---|---|---|---|---|---|
| Sown in Potato | Freshly Harvested | Rest | Another Crop | ||
| Amarantaceae | Amaranthus dubius | *C – *P | C – P | ||
| Asteraceae | Acmella mutisii | C – P | |||
| Galinsoga quadriradiata | C – P | ||||
| Brassicaceae | Raphanus raphanistrum | C | |||
| Fabaceae | Trifolium repens | P | |||
| Juncaceae | Juncus microcephalus | P | |||
| Oxalidaceae | Oxalis corniculate | P | |||
| Phytolaccaceae | Phytolacca bogotensis | C | |||
| Poaceae | Holcus lanatus | C | |||
| Pennisetum clandestinum | C – P | C – P | C – P | C – P | |
| Poa annua | C | ||||
| Zea mays | C – P | ||||
| Polygonaceae | Polygonum nepalense | C – P | C – P | C – P | |
| Rumex crispus | C – P | C – P | C – P | ||
| Rosaceas | Fragaria x ananassa | P | |||
| Solanaceae | Solanum tuberosum | C – P | C – P | C – P | |
| Verbenaceae | Verbena litoralis | C – P | C – P | ||
| *C (cystosori) and P (plasmodia) |
Table 3: Species that Presented Structures Associated with S. Subterranea in the Municipality of La Unión (Antioquia), Evaluated
| Family | Species | Lot Type | ||
|---|---|---|---|---|
| Sown in Potato | Freshly Harvested | Rest | ||
| Asteraceae | Bidens pilosa | *C | ||
| Hypochaeris radicata | C | |||
| Sonchus oleraceus | C | |||
| Fabaceae | Trifolium repens | C | ||
| Poaceae | Pennisetum clandestinum | C – *P | C – P | C – P |
| Polygonaceae | Polygonum nepalense | C | ||
| Solanaceae | Solanum tuberosum | P | P | P |
| *C (Cystosori) And P (Plasmodia) | ||||
| C | P | |||
| A. A. mutissii | B. B. pilosa | C. Fragaria x ananassa | D. G. quadriradiata | |
| C | P | P | C | |
| E. Holcus lanatus | F. J. microcephalus | G. O. corniculata | H. P. clandestinum | |
| C | C | P | C | |
| I. S. tuberosum | I. P. bogotensis | K. V. litoralis | L. Zea mays |
Table 4: Species That Presented Structures Associated with S. Subterranea in the Municipality of Santa Rosa De Osos (Antioquia),
The species exhibiting structures associated with the pathogen are classified within the families Amarantaceae, Asteraceae, Brassicaseae, Fabaceae, Poaceae, Polygonaceae, and Solanaceae. These families have previously been documented as hosts of S. subterranean [12, 16, 18, 29]. Additionally, this study confirms the presence of pathogen structures in the families Juncaceae, Oxalidaceae, Phytolaccaceae, Rosaceae, and Verbenaceae, with the species J. microcephalus, O. cornuculata, P. bogotensis, Fragaria x ananassa, and V. litoralis. These species had previously been reported as hosts of S. subterranea [30].
The galls formation of S. subterranea in Solanum nigrum. They discovered that 83% of potato plants and 52% of S. nigrum exhibited inoculum gills from potatoes. Furthermore, 10% and 31% of potato and S. nigrum plants, respectively, displayed gills when inoculated with gills from S. nigrum. This finding suggests that S. subterranea can complete its life cycle in species other than potatoes and serve as a significant source of inoculum for the spread of the disease [31].
Other study by inoculating three tomato cultivars, “Grape,” “Rome,” and “Truss,” with S. subterranea. They observed a significant decrease in shoot length and fresh weight of the plants, concluding that, similar to potatoes, root infection by this pathogen can hinder the growth of tomato plants and potentially impact the production performance of other host species [5].
Reported Papaver somniferum and Tanacetum cinerariifolium as hosts for S. subterranean [32]. A study in Israel by Tsror, et al. [14] reported 20 species hosting S. subterranea, including two cultivable species (Arachis hypogaea and Triticum aestivum) and 18 weeds (Amaranthus albus, Ammi majus, Astragalus hauraensis, Chenopodium murale, Chenopodium opulifolium, Chrysanthemum segetum, Cynodon dactylon, Malva nicaeensis, Medicago sativa, Phalaris minor, Phalaris paradoxa, Rostaria cristata, Salsola soda, Setaria verticillata, Sinapis nigra, Solanum elaeagnifolium, Tribulus terrestris, and Verbesina encelioides). These weeds were reported for the first time as hosts in this study.
In batches of potatoes, a higher number of species with structures in their roots associated with S. subterranea were found, followed by freshly harvested batches, resting batches, and other crops, as indicated in Tables 3 and 4. This suggests that in batches with potato cultivation, the pathogen is more active. In contrast, in other batches, some species may act as host plants where the pathogen fails to complete its cycle [33, 34]. Only zoosporangians were found in the roots of plants in these batches. Arcila Aristizábal IM, et al. [30] discovered that in controlled conditions, species such as Cyphomandra betacea, Physalis peruvianum, Solanum nigrum, Allium cepa, Solanum quitoense, and Rumex crispus reduce the presence of cystosori and zoosporangia after being planted in three consecutive plantings.
Based on the observed structures, the species are categorized as follows: S. oleraceus, B. pilosa, T. repens, H. radicata, P. annua, R. raphanistrum, P. bogotensis, and H. lanatus only presented cystosori; O. corniculata, J. microcephalus, Fragaria x ananassa, and T. repens only presented plasmodia formation, and P. nepalense, P. clandestinum, S. tuberosum, A. dubius, V. litoralis, R. crispus, G. quadriradiata, Z. mays, and A. mutisii presented both cystosoros and plasmodia within the roots. Observed the presence of zoosporangia and sporeballs of S. subterranea in the species B. Pilosa [16].
Among the species in which structures associated with S. subterranea were found are two crop species Zea mays (maize), where it was possible to observe cystosori and plasmodia, and Fragaria x ananassa (strawberry), where plasmodia were observed (Figure 1G & 1M). That the species Allium cepa, Solanum lycopersicum, Zea mays, Avena sativa, and Triticum aestivum were hosts for both zoosporangia and sporeballs, while Phaseolus vulgaris and Brassica juncea were hosts for sporosorial only [16]. Was evaluated Z. mays in artificial inoculations under hydroponic cultures without showing the presence of structures, that this species is not a host of P. graminis and P. betae [35, 36]. However, weed plant species of the genus Zea are reported as hosts of P. graminis [37].
Structures associated with the pathogen were identified in weed plants such as H. radicata, G. quadriradiata, A. mutisii, J. microcephalus, P. clandestinum, P. annua, and H. lanatus, which have not been previously reported as hosts of S. subterranea. P. annua was previously assessed Andersen BAB, et al. [17] with no observed structures. Additionally, structures were observed in A. dubius, R. raphanistrum, P. Nepalese, R. crispus, S. oleraceus, and T. repens, which have not been reported as hosts of S. subterranea. However, other species within the same genus, such as A. retroflexus, R. sativus, and T. pretense [18], as well as P. avicular, P. convolvulus, S. arvensis, and R. acetosella Andersen BAB, et al. [17] were reported as hosts. R. acetosella, collected in La Unión, did not show pathogen structures upon microscopic analysis.
Weed plants like B. pilosa, S. oleraceus, H. radicata, A. mutisii, R. raphanistrum, T. repens, J. microcephalus, O. corniculata, P. bogotensis, P. clandestinum, H. lanatus, R. crispus, and V. litoralis, exhibited structures associated with S. subterranea, but there are no prior reports of them hosting other plasmodiophoromycetes. Similarly, structures were observed in A. dubius, P. annua, P. Nepalese, and G. quadriradiata, where there are no reports of them being hosts for S. subterranea or other plasmodiophoromycetes. Nonetheless, studies have been conducted on species within these same genera, such as A. reflexus, P. compresa, P. pratensis, P. persicaria Barr DJS , et al. [35], and G. parviflora [38], without any documented structures associated with plasmodiophoromycetes.
The high incidence and severity of the disease are directly linked to increased pathogen inoculum levels. A study assessing the relationship between potato powdery scab inoculum and host resistance found significantly higher concentrations in plots with elevated inoculum levels [4], underscoring the importance of controlling species that facilitate the completion of the pathogen cycle. Establishing the host range of S. subterranea is crucial to fully understand the epidemiology, pathogenesis and management measures of this pathogen [39].
PCR Detection of S. subterranea and Other Plasmodiophoromycetes
Following the microscopic identification of structures associated with S. subterranea, a molecular analysis was conducted due to the structural similarities with other Plasmodiophoromycetes. The molecular analysis involved the utilization of primers for detecting various plasmodiophoromycetes. A positive control for S. subterranea DNA was sourced from potato galls in the municipality of La Unión, while DNA from a cabbage sample exhibiting disease symptoms served as the positive control for P. brassicae. A total of 59 samples, accounting for 5.42% of the overall samples collected, underwent molecular testing.
S. subterranea was successfully identified via PCR using the primer pairs SSR–SSF [26] and Spo8–Spo9 [22]. The species S. tuberosum, present in potato plots in Santa Rosa (Figure. 3), and a freshly harvested plot in Santa Rosa, where cystosori were observed (Figure 1B), both tested positive for the pathogen.
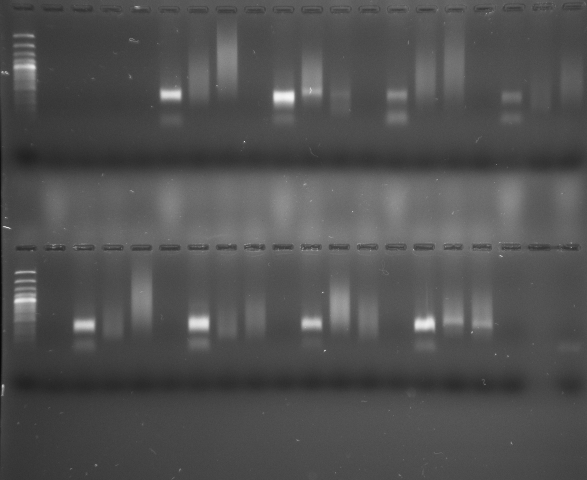
The presence of P. betae was confirmed through PCR analysis, employing the PNS1-BET1 primers. Positive detection occurred in S. tuberosum plants obtained from a freshly harvested potato plot in the municipality of Santa Rosa. Furthermore, P. betae was identified in O. corniculata plants within a lot cultivated with potatoes in the municipality of La Unión, and in T. repens plants in another plot within the same municipality. Notably, Z. mays in the municipality of La Unión, situated in a freshly harvested plot, also exhibited positive results for P. betae (Figures 4 & 5).
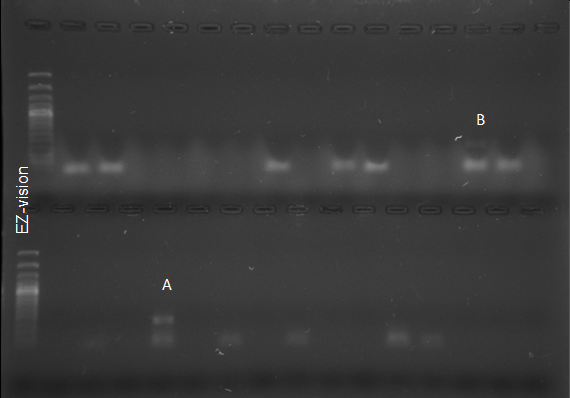
PCR analysis, utilizing the primers ITS1-GRA2 and PNS1- GRA2, confirmed the presence of P_. graminis_ in Z. mays plants within a freshly harvested plot in the municipality of La Unión, where both cystosori and plasmodia were observed (Figures 3 & 5). These findings diverge from those of Barr K, et al. [40] who, through morphological analysis, concluded that maize did not act as a host for P. graminis.
The absence of detection for S. subterranea f. sp. subterranea in some samples could be attributed to the limited set of primers (SPo8, SPo9, SSF, and SSR) used, as they might not be comprehensive enough to identify all the variants of S. subterranea present in Colombia. This limitation is notable, especially given that the identified cystosori structures closely resemble those found in potato galls formed by S. subterranean [41].
Interestingly, the molecular analysis did not yield any positive detection for S. subterranea or the studied plasmodiophoromycetes in the remaining samples. P. brassicae was not identified in any sample. Nevertheless, positive control reactions consistently produced amplicons in all cases, indicating the appropriateness of the extraction and amplification methodologies (Figure 6).
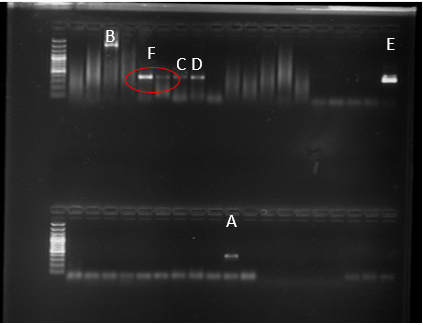
Figure 6: Fragment Amplified for P. Graminis with the Primer ITS1-GRA2 (A), PNS2-GRA2 (B) from Z. Mays. Fragment Amplified For P. Betae with the Primers PNS1 - BET1 from O. Curniculata (C) And from Z. Mays (D). Amplified Fragment for the Sample Used as a Positive Control Of P.Brassicae. (E), Amplified Fragment of P. Betae With PNS1 - BET1 Primers in S. Tuberosum Plants (F).
The observed disparity between structures identified through microscopic analysis and positive PCR samples may be attributed, in part, to the potential presence of PCR inhibitors within the plants. Inhibitors such as phenolic compounds and elevated DNA concentrations [5, 42, 43], could influence the PCR outcomes. Despite employing two different DNA concentrations (1 µL DNA and a 1:100 dilution of DNA), no significant differences in results were obtained. Additionally, it’s worth noting that these primers were initially designed and optimized for potato samples. Consequently, when applied across 48 diverse plant species in this study, variations in amplification efficiency may occur.
Effective crop rotation disrupts the lifecycle of the pathogen, reducing the inoculum levels in the soil and minimizing disease incidence in subsequent potato crops. One effective rotation involves alternating potatoes with cereals different from corn, which are non-host crops for S. subterranea. These crops can help break the disease cycle by preventing the buildup of pathogen inoculum. Another successful rotation includes legumes like beans or peas, which improve soil health and structure, further hindering the survival of the pathogen. Incorporating cover crops such radish, known for their biofumigation properties, can also suppress the pathogen’s presence in the soil and the planting of resistant potato varieties [39]. By integrating these diverse crops into a rotation schedule, farmers can significantly mitigate the impact of powdery scab and sustain healthier potato yields.
Conclusions
Among the forty-eight plant species evaluated, seventeen exhibited structures associated with S. subterranea in La Unión municipality, and seven in Santa Rosa. Notably, none of the evaluated species displayed symptoms of root galls. The identified species with observed structures belong to diverse plant families, including Amarantaceae, Asteraceae, Brassicaceae, Fabaceae, Juncaceae, Oxalidaceae, Phytolaccaceae, Poaceae, Polygonaceae, Rosaceae, Solanaceae, and Verbenaceae. In the case of cultivated plant species, Zea mays (maize) and Fragaria x ananassa (strawberry) were found to harbor structures associated with S. subterranea within their roots, signifying their status as hosts for the pathogen. Additionally, various weed species, including B. pilosa, S. oleraceus, H. radicata, A. mutisii, R. raphanistrum, T. repens, P. clandestinum, H. lanatus, R. crispus, A. dubius, P. annua, P. nepalense, and G. quadriradiata, exhibited pathogen structures, despite lacking previous reports of being hosts for other plasmodia. PCR analysis confirmed the presence of S. subterranea in S. tuberosum, P. betae in S. tuberosum, O. corniculata, T. repens, and Z. mays, and P. graminis in Z. mays. Notably, P. brassicae was not detected in any of the analyzed samples. These findings contribute valuable insights into the diversity of host plants for S. subterranea, emphasizing the importance of both cultivated and weed species in the epidemiology of potato powder scab. seeing the importance of developing varieties of Solanum spp resistant to powdery scab, thus contributing to better production and higher quality.
Acknowledgment
The Politécnico Colombiano Jaime Isaza Cadavid, research group “Sistemas Agrícolas Tropicales” (COL0007149) and the Universidad Nacional de Colombia Sede Medellín research group “ Mejoramiento y Producción de Especies Andinas y Tropicales” (COL0039484) and the Federación de Productores de Papa de Colombia (FEDEPAPA).
Funding
This research was financed through the project “ Uso de plantas trampa para la reducción de inóculo de Spongospora subterranea como medida de manejo de la sarna polvosa de la papa y determinación de la susceptibilidad de cultivos de rotación “ co-financed by the Ministerio de Agricultura y Desarrollo Rural (MADR), Asociación Hortofrutícola de Colombia (ASOHOFRUCOL).
Conflicts of Interest
The authors declare no conflict of interest
References
-
Mallik I, Fulladolsa AC, Yellareddygari SKR, Bittara FG, Charkowski AO, et al. (2019) Detection and quantification of _Spongospora subterranea_ sporosori in soil by quantitative Real-Time PCR. Plant Disease 103(12): 3189-3198.
-
Liu L, Qin L, Zhou Z, Hendriks WGHM, Liu S, et al. (2020) Refining the Life Cycle of _Plasmodiophora brassicae_. Phytopathology 110(10): 1704-1712.
-
Merz U (2008) Powdery scab of potato—occurrence, life cycle and Epidemiology. Am J Pot Res 85(4): 241-246.
-
Brierley JL, Sullivan L, Wale SJ, Hilton AJ, Kiezebrink DT, et al. (2013) Relationship between _Spongospora_ _subterranea_ f. sp. _subterranea_ soil inoculum level, host resistance and powdery scab on potato tubers in the field. Plant Pathology 62(2): 413-420.
-
Balendres MA, Tegg RS, Amponsah J, Wilson CR (2018) Zoosporangial root infection of tomato by _Spongospora_ _subterranea_ in hydroponic and glasshouse culture results in diminished plant growth. Journal of Phytopathology 166(6): 412-419.
-
Simango K, Van der Waals JE (2017) Effects of Different Soil Treatments on the Development of _Spongospora_ _subterranea_ f. sp. _subterranea_ in Potato Roots and Tubers in the Greenhouse. Potato Res 60(1): 47-60.
-
Brierley JL, Stewart JA, Lees AK (2009) Quantifying potato pathogen DNA in soil. Applied Soil Ecology 41(2): 234-238.
-
Balendres M, Cumagun C, Woodhall J, Whitworth J (2024) Powdery Scab of Potato: A Diagnostic Guide. Powdery Scab of Potato: A Diagnostic Guide pp: 22.
-
Hernandez ML, Falloon RE, Butler RC, Conner AJ, Bulman SR (2013) _Spongospora subterranea_ root infection assessed in two potato cultivars differing in susceptibility to tuber powdery scab. Plant Pathology 62(5): 1089-1096.
-
Sparrow LA, Rettke M, Corkrey SR (2015) Eight years of annual monitoring of DNA of soil-borne potato pathogens in farm soils in south eastern Australia. Australasian Plant Pathol 44(2): 191-203.
-
Falloon RE, Merz U, Butler RC, Curtin D, Lister RA, et al. (2016) Root infection of potato by _Spongospora_ _subterranea_: knowledge review and evidence for decreased plant productivity. Plant Pathology 65(3): 422-434.
-
Neuhauser S, Kirchmair M, Bulman S, Bass D (2014) Cross-kingdom host shifts of phytomyxid parasites. BMC Evolutionary Biology 4(1): 33.
-
Abbas A, Madadi M (2016) A review paper on potato mop-top virus (PMTV): Occurrence, properties and management. World Journal of Biology and Biotechnology 1(3):129-134.
-
Tsror (Lahkim) L, Erlich O, Hazanovsky M, Lebiush S (2019) Control of potato powdery scab (_Spongospora_ _subterranea_) in Israel with chloropicrin, metam sodium or fluazinam. Crop Protection. 124: 104836.
-
Qu X, Christ BJ (2004) Genetic variation and phylogeny of _Spongospora subterranea_ f.sp. _subterranea_ based on ribosomal DNA sequence analysis. Am J Pot Res 81(6): 385-394.
-
Simango K, Slabbert CP, Van der Waals JE (2020) Alternative hosts of _Spongospora subterranea_ f. sp. _subterranea_ in southern Africa. Eur J Plant Pathol 157(2): 421-424.
-
Andersen BAB, Nicolaisen M, Nielsen SL (2002) Alternative hosts for potato mop-top virus, genus Pomovirus and its vector _Spongospora subterranea_ f.sp. _subterranea_. Potato Res 45(1): 37-43.
-
Qu X, Christ BJ (2006) The host range of _Spongospora_ _subterranea_ f. sp. _subterranea_ in the United States. Am J Pot Res 83(4): 343-347.
-
Merz U (1997) Microscopical observations of the primary zoospores of _Spongospora subterranea_ f.sp. _subterranea_. Plant Pathology 46(5): 670-674.
-
Montero-Astúa M, Vásquez V, Rivera C (2004) Potato powdery scab, caused by _Spongospora subterranea_ f. sp. _subterranea_ in Costa Rica. Potato Res 47(1): 25-34.
-
Merz U, Walsh JA, Bouchek-Mechiche K, Oberhänsli Th, Bitterlin W (2005) Improved immunological detection of _Spongospora subterranea_. Eur J Plant Pathol 111(4): 371-379.
-
Bulman SR, Marshall JW (1998) Detection of _Spongospora_ _subterranea_ in potato tuber lesions using the polymerase chain reaction (PCR). Plant Pathology 47(6): 759-766.
-
Qu XS, Wanner LA, Christ BJ (2011) Multiplex real‐time PCR (TaqMan) assay for the simultaneous detection and discrimination of potato powdery and common scab diseases and pathogens. Journal of Applied Microbiology 110(3): 769-777.
-
O’Brien PA, Milroy SP (2017) Towards biological control of _Spongospora subterranea_ f. sp. _subterranea_, the causal agent of powdery scab in potato. Australasian Plant Pathol 46(1): 1-10.
-
Irmayanti S, Winarni I (2022) Modification of root staining preparation method for detection and visualization colony formation of arbuscular mycorhriza fungi (AMF). MANILKARA: Journal of Bioscience 1(1): 09-18.
-
Qu X, Kavanagh J, Egan D (1998) Development of specific and sensitive primers for the identification of _Spongospora subterranea_ f. sp. _subterranea_. Research report 1996-1997, faculty of agriculture. pp: 206-207.
-
Nickrent DL, Schuette KP, Starr EM (1994) A molecular phylogeny of Arceuthobium (Viscaceae) based on nuclear ribosomal DNA internal transcribed spacer sequences. American Journal of Botany 81(9): 1149-1160.
-
W White T, Bruns T, Lee S and Taylor J (1990) Chapter - Amplification and direct sequencing of the fungal ribosomal rna genes for phylogenetics. In: Innis MA, Gelfand DH, et al. (Eds.), PCR protocols. A guide to methods and applications. Academic Press, New York. 482p.
-
Tsror (Lahkim) L, Shapira R, Erlich O, Hazanovsky M, Lebiush S (2020) Characterization of weeds and rotational crops as alternative hosts of _Spongospora_ _subterranea_, the causal agent of powdery scab in Israel. Plant Pathology 69(2): 294-301.
-
Arcila Aristizábal IM, González Jaimes EP, Zuluaga Amaya CM, Marín Montoya MA, Cotes Torres JM (2014) Effect of _Spongospora subterranea_ f. sp. _subterranea_ Infection on Alternate Hosts. Rev Fac. Nac Agron 67(2): 7261-7269.
-
Nitzan N, Boydston R, Batchelor D, Crosslin J, Hamlin L, et al. (2009) Hairy Nightshade is an Alternative Host of _Spongospora subterranea_, the Potato Powdery Scab Pathogen. Am J Pot Res 86(4): 297-303.
-
Clark TJ, Rockliff LA, Tegg RS, Balendres MA, Amponsah J, et al. (2018) Susceptibility of opium poppy and pyrethrum to root infection by _Spongospora subterranea_. Journal of Phytopathology 166(10): 694-700.
-
Aristizabal IMA, Jaimes EPG, Amaya CMZ, Torres JMC (2013) Alternate hosts of _Spongospora subterranea_ f. sp. _subterranea_ identification in Colombia by bioassay. Rev Fac Nac Agron 2(66): 6987-6998.
-
Alaryan MM, Zeng Y, Fulladolsa AC, Charkowski AO (2023) Brassica cover crops and natural _Spongospora_ _subterranea_ infestation of peat-based potting mix may increase powdery scab risk on potato. Plant Disease 107(9): 2769-77.
-
Barr DJS (1979) Morphology and host range of _Polymyxa_ _graminis_, _Polymyxa betae_, and _Ligniera pilorum_ from ontario and some other areas. Canadian Journal of Plant Pathology 1(2): 85-94.
-
Jin Y, Rouse M, Groth J (2014) Population Diversity of _Puccinia graminis_ is sustained through sexual cycle on alternate hosts. Journal of Integrative Agriculture 13(2): 262-264.
-
Suproniene S, Kadziene G, Irzykowski W, Sneideris D, Ivanauskas A, et al. (2019) Weed species within cereal crop rotations can serve as alternative hosts for _Fusarium graminearum_ causing Fusarium head blight of wheat. Fungal Ecology 37: 30-37.
-
Mouhanna AM, Langen G, Schlösser E (2008) Weeds as alternative hosts for BSBV, BNYVV, and the vector _Polymyxa betae_ (German isolate). J Plant Dis Prot 115(5): 193-198.
-
Strydom RF, Wilson CR, Tegg RS, Balendres MA, Van Der Waals JE (2024) Advancements in _Spongospora_ _subterranea_: Current Knowledge, Management Strategies, and Research Gaps. European Potato Journal.
-
Barr KJ, Asher MJC (1992) The host range of _Polymyxa_ _betae_ in Britain. Plant Pathology 41(1): 64-68.
-
Pandey B, Mallik I, Gudmestad NC (2020) Development and application of a real-time reverse-transcription PCR and droplet digital PCR Assays for the Direct Detection of Potato mop top virus in Soil. Phytopathology 110(1): 58-67.
-
Daniell TJ, Davidson J, Alexander CJ, Caul S, Roberts DM (2012) Improved real-time PCR estimation of gene copy number in soil extracts using an artificial reference. Journal of Microbiological Methods 91(1): 38-44.
-
Thangavel T, Tegg RS, Wilson CR (2015) Monitoring _Spongospora subterranea_ Development in Potato Roots Reveals Distinct Infection Patterns and Enables Efficient Assessment of Disease Control Methods. PLOS ONE 10(9): 0137647.
- Enhancement of Vegetative Growth and Fruit Yield in Cucumber (Cucumis sativus L.) via Spiritual Blessing (Biofield) Energy Intervention
- Production of Açaí (Euterpe oleracea Mart.) under Different Agroforestry System Management Intensities in Amazonian Floodplain (Varzea) Forests
- Coffee and the Production Region: What is the Secret to the Expression "Quality"?
- Experiential Agripreneurship Training in Sub-Saharan Africa: Integrating a Business Incubator into Postgraduate Livestock Education at the University of Buea
- Advances in Agricultural High-Quality Development
- Linking Compost Residue to ABAGE in Plants - a Short Note