Par-4 Therapy Sensitizes Glioblastoma Cells to Radiotherapy and Chemotherapy
<p>Glioblastoma multiforme (GBM) is the most aggressive and common neurological tumor and presents with a dismal prognosis. Currently treatments to treat GBM include surgery, radiation therapy, and chemotherapy, typically Temozolamide (TMZ). Recurrence, however, is almost inevitable and the median survival for these patients is only 14.6 months. This study explored the potential of sensitizing GBM cells with Prostate apoptosis response protein-4 (Par-4). Par-4 protein expression was up-regulated in three GBM cell lines in vitro followed by treatment with standard radiotherapy and chemotherapy regimens. Par-4 up-regulation in these cell lines increased apoptosis of tumor cells alone and in combination with chemotherapy, radiotherapy, or both. In addition, cells were treated with the phenylalkyl isoselenocyanate, ISC-4, to induce activation of Par-4, and the combined treatment of ISC-4 with Par-4 enhanced apoptosis in the GBM cells. Utilizing a molecular target for GBM could lead to more permanent regression of tumor in addition to increased response to therapies.</p>
Introduction
Glioblastomas are highly malignant brain tumors arising from astrocytes, the supportive tissues of the brain. They reproduce quickly, contain a wide array of cell types, and are supported by a large network of blood vessels, which contributes significantly to their dismal prognosis and necessitates a treatment plan incorporating several approaches [1]. Current treatment includes surgery to debulk the tumor, followed by radiation therapy and chemotherapy, typically TMZ [1].
Surgery is rarely sufficient for removal of all malignant cells, as the primary tumor is extensively locally invasive and has the potential to spread invasively early. Often unknown microscopic lesions exist that have invaded surrounding brain tissue at the time of diagnosis, leaving some unresectable tumor. Chemotherapy has a limited effect as well, partially because of the toxic effects on normal tissue and the blood/brain barrier [2]. TMZ penetrates the blood brain barrier and can be taken orally. In a seminal study Stupp, showed that treating patients with TMZ concomitant with radiotherapy followed by adjuvant TMZ offered better results than radiotherapy alone [3], yet, even increasing the treatment cycles had little effect on survival [4]. However, many glioblastomas are resistant to TMZ, likely resulting from altered DNA methylation by O6-methylguaning methyltransferase [5]. Despite utilizing an aggressive regimen, the 5-year survival rate for GBM is 15-20%, underscoring the need to utilize molecular targets in the treatment of high-grade gliomas. GBM can be either primary, which arises de novo and composes about 90% of the cases, or secondary, which develops from lower grade astrocytoma. Primary GBM is associated with a worse prognosis and displays a slightly different genetic profile than secondary GBM. Common genetic alterations in primary glioblastoma include amplification of oncogenes, commonly EGFR and CDK4, and deletion of tumor suppressor genes, most commonly PTEN, and loss of chromosome 10 [1]. TMZ is an alkylating agent that causes arrest in the G2/M phase of the cell cycle in p53-intact gliomas cells and transient G2/M arrest in p53-inactivated gliomas. Where as it is the standard therapy used for gliomas after surgery and irradiation, often patients present with tumors resistant to TMZ [6, 7], leaving few options. Although TMZ has contributed to the survival of glioma patients, the addition of TMZ to radiotherapy has led only to a longer median survival time, but patients eventually die from their illness [3]. Furthermore, although TMZ causes cell cycle arrest, this delay can also allow for more time to repair the DNA lesions induced by TMZ, and thus decrease its effectiveness [8]. There is a need for targeted molecular therapies not only to halt the cell cycle, but also to induce apoptosis to reduce the development of chemotherapy resistance. Certain cell lines, including A172 and T98G have already shown to have resistance to TMZ. This has been linked to the expression of certain levels of DNA alkylating proteins and repair enzymes that confer resistance to TMZ in these cell lines [5]. Although all cell lines have the potential to benefit from Par-4 molecular therapy, A172 and T98G especially have the potential to gain significant benefit from the use of Par-4 as a targeted molecular therapy. Because current standard of care is sub-optimal, this study investigated the unique combination of increased Par-4 expression with a novel therapy both with and without standard of care therapies. Par-4 is a pro- apoptotic protein, encoded by the human PAWR gene_,_ which activates both intrinsic and extrinsic pathways of programmed cell death selectively in cancer cells [9, 10]. It has been shown to up-regulate apoptosis in cancer cells, including colon and prostate, and has been shown to stimulate apoptosis without additional apoptotic agents [11, 12]. The intrinsic pathway involves mitochondrial- mediated apoptosis [9]. The extrinsic pathway involves the activation of TRAIL. In this case, Par-4 exhibits bystander effects, in that cells expressing Par-4 can secrete the protein which will bind to a glucose related protein, Grp78, on nearby or distal cancer cells that do not over express Par-4 and induce apoptosis [9]. Figure 1 Par- 4 has been shown to sensitize cells to both chemotherapy and radiation therapy [13, 14]. Par-4 is inactivated by Akt1, and, thus, is inactive in many cancers [15]. Par-4 also is down regulated in a number of cancers, including endometrial [16], breast [17, 18], pancreatic [19], and colon [12] cancers. Over expression of Par-4 can increase the sensitivity of many of these cancer cells to apoptotic agents. A potential role for Par-4 in glioma cell sensitivity has been suggested by the finding that inhibition of NFκB reduces the tumorigenicity of glioma stem cells and sensitizes them to TMZ [20], supporting our hypothesis that up regulation and/or activation of Par-4 in GBM cells will sensitize them to radiation therapy and to TMZ. Finally, the fact that Par-4 is secreted to exert its effects on both proximally and distally located cancer cells increases the potential success of the treatment.
![Figure 1: Schematic of effects of Par-4 on cancer cells. Activated Par-4 causes apoptosis through both intrinsic (1) and extrinsic pathways (2) when cells are exposed to apoptotic agents. Akt1 inactivates Par-4 through phosphorylation; however, inhibition of Akt1 with ISC-4 allows Par-4 to remain active Since Par-4 can be present in cells but inactivated by Akt1 via phosphorylation, additional treatment to inhibit Akt1 activity in conjunction with Par-4 molecular therapy has the potential to increase tumor apoptosis. Phenylalkyl isoselenocyanates (ISCs) are selenium derivatives of the isothiocyanate class and have been shown to inhibit Akt activity [21]. ISC-4 is an ISC that was developed and determined to have significant anti-tumor activity in melanoma as a mono-agent _in vivo_ and in colon cancer xenographs as a mono-agent and in combination with 5- Fluorouracil [22]. ISC-4 therapy in the context of glioblastoma is largely unexplored and a synergistic combination with radiation or Par-4 could be a promising therapy.](/fulltextimages/1228/fig_1.png)
Figure 1: Schematic of effects of Par-4 on cancer cells. Activated Par-4 causes apoptosis through both intrinsic (1) and extrinsic pathways (2) when cells are exposed to apoptotic agents. Akt1 inactivates Par-4 through phosphorylation; however, inhibition of Akt1 with ISC-4 allows Par-4 to remain active Since Par-4 can be present in cells but inactivated by Akt1 via phosphorylation, additional treatment to inhibit Akt1 activity in conjunction with Par-4 molecular therapy has the potential to increase tumor apoptosis. Phenylalkyl isoselenocyanates (ISCs) are selenium derivatives of the isothiocyanate class and have been shown to inhibit Akt activity [21]. ISC-4 is an ISC that was developed and determined to have significant anti-tumor activity in melanoma as a mono-agent in vivo and in colon cancer xenographs as a mono-agent and in combination with 5- Fluorouracil [22]. ISC-4 therapy in the context of glioblastoma is largely unexplored and a synergistic combination with radiation or Par-4 could be a promising therapy.
Materials and Methods
Cell Culture
Human GBM cell lines A172, SNB19, and T98G were used for these studies. These cells are available from American Type Culture Collection (ATCC, https://www.atcc.org). Cells were plated at a density of 1.8x105 cells/well in a 6-well tissue culture dish and cultured in RPMI (Cellgro, Manassas, VA) containing 10% FBS at 37° C and 5% CO2. Human colorectal cancer cells SW480 and HT-29 were obtained from ATCC and maintained in RPMI + 10% FBS at 37° C and 5% CO2 and seeded in 6 well plates at a density of 300,000 cells/well.
Transfection
All cell lines were transfected transiently with Par-4 cDNA using Signa Gen Polyjet (SignaGen Laboratories, Rockville, MD), according to the manufacturer's instructions at different dosing schemes to determine the most efficient transfection scheme, as well as the one that worked the best with chemotherapy and/or radiotherapy. Plasmid DNA, consisting of a eukaryotic expression plasmid containing the human PAWR gene (Ori Gene, Rockville, MD), was combined with Polyjet at a ratio of 1:2, w/v, incubated for 5 minutes, and added to the cells at a concentration of 0. 5 μg/ml. Control cells were treated with the transfection agent alone or empty plasmid to ensure transfection efficiency and non-toxicity of the liposomes and base plasmid. Cells were harvested after 24, 48, and 72 hours and assessed for Par-4 expression by Western blot analysis. For conditioned media experiments, both mock (empty plasmid) and Par- 4 stably transfected SNB19 cells were grown to 80% confluence, a stage at which cells were not undergoing significant apoptosis in either condition. The media was removed, cells were washed in PBS, and fresh media was added and incubated for 8 hours. This conditioned media was then removed and added to wild type cells 12 hours prior to additional treatment. Further treatment was performed for 48 hours.
Western Blotting
Cells were treated in log-phase growth, harvested by cell scraping in RIPA buffer containing protease inhibitors, centrifuged, and lysed on ice for 30 minutes. The supernatant was collected following centrifugation, and protein concentration was determined using the Pierce BCA Protein Assay (Rockford, IL). Samples were electrophoresed by SDS-PAGE on 10% Tris gels, transferred to nitrocellulose membranes (BioRad), and blocked in 5% non-fat milk in TBST for 1 hour. Membranes then were incubated with primary antibodies Par-4 and Grp78 (sc 1666 and sc13968, respectively, Santa Cruz Biotechnology, Dallas, TX) and Akt1, caspase 3, pPar-4 (2967, 9662, and 2329, respectively, Cell Signaling, Danvers, MA) at 1:1000 in 5% non-fat milk in TBST overnight at 4°C. Membranes were washed in TBST, incubated with the appropriate HRP-conjugated secondary antibody (Thermo-Scientific) for 1 hour, washed in TBST, developed using ECL-Plus, and visualized by exposure to X-Ray film (Thermo-Scientific).
Treatment with TMZ and ISC-4
SNB19, A172, and T98G cells were treated with increasing concentrations as indicated in each figure of TMZ and ISC-4 suspended in DMSO (total DMSO concentration in solution 0.0025%) for 48 hours to determine EC50. Control cells were treated with DMSO.
Radiation and MTT assay
Cells were plated in 6-well plates at a density of 1.8x105 cells/well and transiently transfected with a Par- 4 expression plasmid. The cells then were exposed to radiation doses of 1 Gy, 2 Gy, 4 Gy, 8 Gy, 16 Gy, 32 Gy, and 64 Gy using a Cobalt 60 source emitting 1.25 MV gamma rays. The schedule of dosing, including frequency and number of treatments, was determined and calculated empirically. Efficacy of treatment was assessed using a 3- (4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay according to the manufacturer’s instructions (Calbiochem) to determine metabolic activity 24-72 hours after treatment. Results were assessed by measuring the absorbance at 570 nm, with 630 nm absorbance as the reference. Percent viability is expressed as the absorbance normalized to the DMSO control.
Conditioned Medium Treatment
Wild type cells were plated in 12 well plates. Cells transected with either Par-4 or empty vector (Mock) were grown to 80% confluence, at which time cells were not apoptotic. Media was removed and cells were washed. Fresh media was added and allowed to incubate for 8 hours to create conditioned media. Wild type cells were washed, the conditioned media was added, and cells were incubated for 12 hours after which ISC-4 and TMZ were added for an additional 48 hours. Statistical analysis was performed using 2-way ANOVA for comparison of all treatment options, as the response to radiation and ISC-4 can be affected by whether the cells over expressed Par-4 or not.
Results
Investigation of Par-4 Expression in Glioblastoma Cells
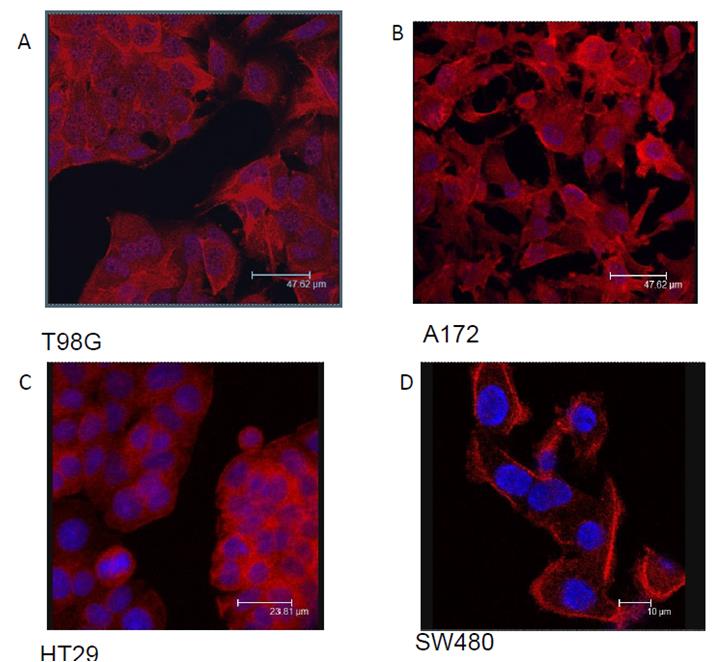
Three glioblastoma cell lines with varied genotypes as found from ATCC were selected to conduct these studies. The A172 genotype contains phosphatase and tensin homolog (PTEN) and CDKN2A mutations. T98G, while also associated with CDK2A mutations, lacks p53 activity, which would normally arrest cell cycle for DNA damage, induce apoptosis, and activate expression of diverse set of genes. SNB19 is known to be particularly aggressive, as these tumors have been associated with mutations in both PTEN and p53 and this combined loss of function can result in increased tumorigenicity. The SNB19 cell line has been utilized in a wide range of studies related to the basic properties of human glioblastoma multiforme, with well-studied immunologic, biochemical, and cytogenic properties [23, 24]. Along with A172 and T98G cell lines, the SNB19 cell line has important utility in the investigation and pathophysiology of human malignant gliomas. Par-4 has been shown to translocate to the nucleus upon activation [25]. Since Par-4 was shown to be present in GBM cells, we explored the subcellular location with particular interest in the nucleus of the cells. Fluorescence microscopy was performed to determine the presence and subcellular location of Par-4 in GBM cells. In other solid tumors tested, specifically colon cancer cell lines, Par-4 remains in the cytoplasm until activated. Results indicate that Par-4 is located throughout the GBM cell lines A172 and T98G, including the nucleus (Figures 2A and B), as opposed to colorectal cancer cells, HT29 and SW480 in which Par-4 is mainly cytoplasmic (Figures 2C and D).
Protein Expression in GBM Cells
GBM cells were examined for endogenous expression of Par-4, Akt 1, and GRP78. Since the presence of GRP78 is instrumental in the ability of extracellular Par-4 to induce apoptosis in cancer cells, the three glioblastoma cell lines were examined for the presence of GRP78 by Western blot analysis. GRP78 was expressed in all three lines, as was Par-4 (Figure 3A). Cells were treated with ISC-4, or vehicle, and assayed for active Par-4 (pPar-4), total Akt1, There was little effect of ISC-4 treatment on Par-4 inT98G cells.
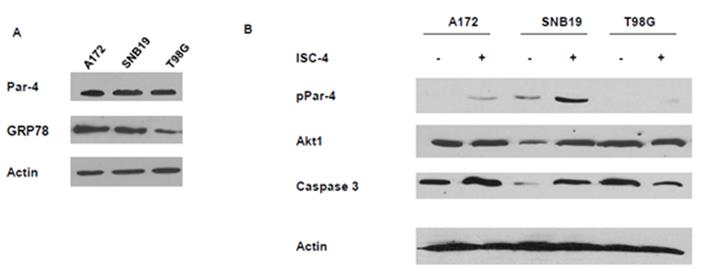
Figure 3: Western blots showing the levels of proteins in GBM cell both treated and untreated with ISC- 4. A. Native levels of Par-4, phospho-Par-4 (active), and GRP 78 in cells. B. Cells were treated with 12.5 μg.ml ISC-4 for 48 hours and whole cell lysates made. Panel B shows the levels of phospho-Par-4, Akt1, and Caspase 3 in three GMB cell lines.
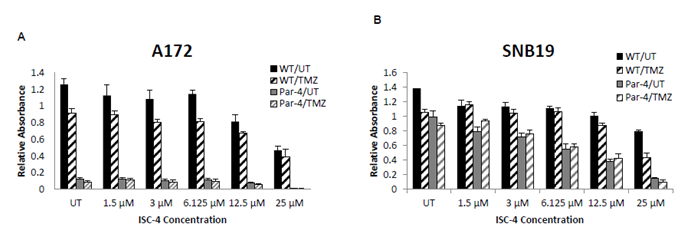
GBM Response to ISC-4 and TMZ therapies
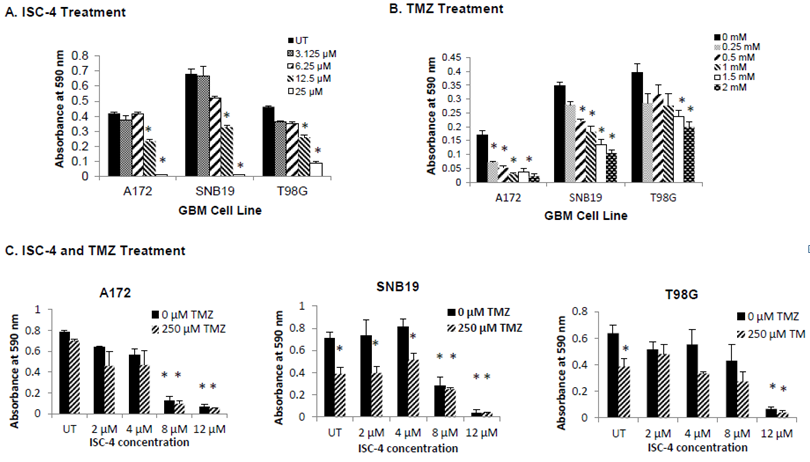
ISC-4 and TMZ were tested in vitro at increasing concentrations to determine the EC50, which was to be used for Par-4 transfection and flow cytometry assays. Glioblastoma cells were sensitive to ISC-4 and TMZ. While each cell line was sensitive to both therapies in a dose-
Determination of Radiation Dose for the SNB19, T98G, and A172 Cell Lines
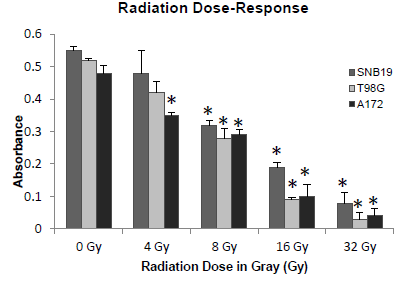
Cells were plated as described and allowed to grow for 24 hours. Cell viability was examined 48 hours post radiation by MTT assay. The sensitivity of the cell lines to radiation treatment was assessed and EC50 was determined to be 10 Gray as portrayed in the radiation exposure curve in (Figure 5) For following experiments, 8 Gray was used in combination with Par-4 treatment for apoptosis assay by flow cytometry.
A172 and SNB19 cell lines were transfected with either mock vector or Par-4 expressing vector at a concentration of 0.5 μg/ml and allowed to grow for 24 hours. Cells were
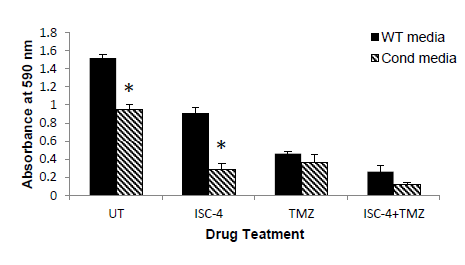
Par-4 is excreted from cells and has been shown to exhibit effects on distal cancer cells through binding of the with chemotherapy. Media was removed from cells transfected with Par-4 or empty plasmid and placed on fresh cells. The cells were incubated for 12 hours, after which cells were treated with 12.5 μM ISC-4, 1.0 mM TMZ, or both. As seen in (Figure 7), when glioblastoma cells were plated in Par-4 conditioned media, ISC-4 showed significantly increased effect, although TMZ did not. This is to be expected as TMZ inhibits cell cycle progression, but may not cause cell death.
Discussion
Par-4 has the potential to be a molecular target for cancer therapies because of its significance in the activation of intrinsic apoptotic pathways. We showed that Par-4 upregulation in glioblastoma cells can induce apoptosis in both the presence and absence of standard chemotherapy drugs. Although the current mainstay of treatment for GBM is a combination of surgery, radiation, and/or chemotherapy, recurrence rates are high and median survival remains low, despite aggressive approaches. We found that simply transfecting Par-4 into glioblastoma cell lines had a significant effect on tumor cell survival, especially in A172 cell lines. This finding is corroborated by Jagtap [26]. The cell lines chosen represent a variety of glioblastoma cell types, with different mutations and molecular markers, displaying a wide range of properties. Furthermore, the combination of standard chemotherapy of TMZ and ISC-4 with upregulation of Par-4 showed a significant increase in apoptosis of GBM cells, suggesting that Par-4 heightens the cell sensitivity to the apoptotic stimulus of the chemotherapy drugs. Radiation alone did not lead to a significant rate of apoptosis; however radiation combined with Par-4 transfection led to a significant increase in apoptosis in all cell lines. Radiation is part of the standard of care for GBM, often in combination with chemotherapy, and these results show that radiation alone did not confer a significant increase in apoptosis compared to control. However, Par-4 and Par-4 plus chemotherapy can significantly increase apoptosis when combined with the same dosage of radiation. This demonstrates the impact of transfection with Par-4, which could be partially reflective of A172’s known susceptibility to resistance of TMZ. It is possible that Par-4 transfection allowed the A172 cell line to be more susceptible to chemotherapy, when otherwise it was partially resistant, but further tests would be needed to determine the specific molecular attributes of this particular A172 line and how it interacted with the TMZ. This has implications for other cell lines that have become resistant to drugs, including A172 and T98G among others, which are known to carry higher risk of resistance to chemotherapy. Also in the A172 cell line, transfection with Par-4 plus radiation was superior to treatment with ISC-4 and radiation. Similar results were seen with T98G cell line, with Par-4 transfection increasing apoptosis when compared to ISC-4 alone. Transfection with Par-4 and radiation without any chemotherapy significantly increased apoptosis in all cell lines, demonstrating its potential to be incorporated as a targeted therapy. Treatment with Par-4 and radiation was significantly better than chemotherapy and radiation in both A172 and T98G cell lines. This shows the potential of Par-4 as a targeted molecular mechanism, because when combined with radiation, Par-4 is superior to standard chemotherapy and radiation in A172 and T98G cell lines. Even in SNB19, a particularly invasive cell line without documented resistance to TMZ, Par-4 transfection significantly increased apoptosis, both with and without added chemotherapy. This indicates that Par-4 upregulates apoptosis in cell lines with a multitude of different properties, not only those with the susceptibility to be resistant to TMZ. Importantly, when GBM cells were plated in Par-4 conditioned media, the combination of ISC-4 and Par-4 significantly induced apoptosis, supporting the presence of an exogenous mechanism of Par-4 sensitization. Par-4 has been shown to cause apoptosis in cancers cells, but specifically we examined Par-4 expression in glioblastoma cells. Par-4 has been shown to translocate to the nucleus upon activation [11]. However, we found that Par-4 is located throughout GBM cells as opposed to colorectal cells. As nuclear localization indicates that Par- 4 is active and inducing apoptosis, and GBM cells are not rapidly undergoing apoptosis, there is likely some additional method of inactivation of Par-4 to prevent this, or perhaps there is a critical concentration of Par-4 needed in the nucleus. In Par-4 sensitized media, ISC-4
and Par-4 significantly induced apoptosis, but TMZ did not. This is to be expected as TMZ inhibits cell cycle progression, but may not cause cell death. ISC-4 was associated with decreased levels of p-Akt in the SNB19 cell line, indicating a mechanism for higher levels of activated Par-4 with ISC-4 treatment. This suggests that the inhibition of Akt with ISC-4 results in activation of Par-4, especially in A172 and SNB19 cell lines. We demonstrate that the presence of phosphorylated Akt is correlated with little to no level of phosphorylated/activated Par-4, which is a potential mechanism in which Akt1 prevents activation of Par-4 in glioblastoma cells, and could be a mechanism of cancer cell avoidance of apoptosis. Additionally, ISC-4 increased levels of GRP78 protein in SNB19 cells (data not shown), another possible mechanism for the increased activity of Par-4. The untreated A172 cell line has high levels of GRP78, which could be a potential mechanism for its sensitivity to Par-4 transfection alone. Due to the high levels of phosphorylated/activated Par-4 with high levels of GRP78, and the role GRP78 plays in the extrinsic apoptosis pathway, there is a possibility that GRP78 plays a role in the mechanism of apoptosis for glioblastoma cells, but would have to be further investigated. Par-4 may play a significant role in the treatment of GBM by increasing the sensitivity of GBM cells to chemo- and radiotherapies to undergo apoptosis. In addition to simple transfection of the A172 cell line leading to significant increases in apoptosis, with all cell lines, the combination of chemotherapy and Par-4 transfection or radiation plus chemotherapy plus Par-4 transfection significantly increased apoptosis when compared to chemotherapy or radiation alone. Our data shows that Par-4, particularly in combination with ISC-4, is a potential molecular target for glioblastoma patients. Our future studies include animal models of GBM and the potential involvement of additional drugs.
References
-
Wilson TA, Karajannis MA, Harter DH (2014) Glioblastoma multiforme: State of the art and future therapeutics. Surg Neurol Int 5: 64.
-
Ung N, Yang I (2015) Nanotechnology to augment immunotherapy for the treatment of glioblastoma multiforme. Journal of neuro-oncology 123(3): 473- 481.
-
Stupp R, Hegi ME, Mason WP, van den Bent MJ, Taphoorn MJ, et al. (2009) Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. The Lancet. Oncology 10(5): 459- 466.
-
Blumenthal DT, Gorlia T, Gilbert MR, Kim MM, Burt Nabors L, et al. (2017) Is more better? The impact of extended adjuvant temozolomide in newly diagnosed glioblastoma: a secondary analysis of EORTC and NRG Oncology/RTOG. Neuro-oncology 19(8): 1119-1126.
-
Lee SY (2016) Temozolomide resistance in glioblastoma multiforme. Genes & Diseases 3:198- 210.
-
Friedman HS, McLendon RE, Kerby T, Dugan M, Bigner SH, et al. (1998) DNA mismatch repair and O6- alkylguanine-DNA alkyltransferase analysis and response to Temodal in newly diagnosed malignant glioma. J Clin Oncol 16(12): 3851-3857.
-
Hegi ME, Liu L, Herman JG, Stupp R, Wick W, et al. (2008) Correlation of O6-methylguanine methyltransferase (MGMT) promoter methylation with clinical outcomes in glioblastoma and clinical strategies to modulate MGMT activity. J Clin Oncol 26(25): 4189-4199.
-
Zhuang D, Liu Y, Mao Y, Gao L, Zhang H, et al. (2012) TMZ-induced PrPc/par-4 interaction promotes the survival of human glioma cells. Int J Cancer 130(2): 309-318.
-
Burikhanov R, Zhao Y, Goswami A, Qiu S, Schwarze SR, et al. (2009) The tumor suppressor Par-4 activates an extrinsic pathway for apoptosis. Cell 138(2): 377-388.
-
Chakraborty M, Qiu SG, Vasudevan KM, Rangnekar VM (2001) Par-4 drives trafficking and activation of Fas and Fasl to induce prostate cancer cell apoptosis and tumor regression. Cancer research 61(19): 7255- 7263.
-
Sells SF, Han SS, Muthukkumar S, Maddiwar N, Johnstone R, et al. (1997) Expression and function of the leucine zipper protein Par-4 in apoptosis. Mol Cell Biol 17(7): 3823-3832.
-
Wang BD, Kline CL, Pastor DM, Olson TL, Frank B, et al. (2010) Prostate apoptosis response protein 4 sensitizes human colon cancer cells to chemotherapeutic 5-FU through mediation of an NF kappaB and microRNA network. Mol Cancer 9: 98.
-
Chendil D, Das A, Dey S, Mohiuddin M, Ahmed MM (2002) Par-4, a pro-apoptotic gene, inhibits radiation- induced NF kappa B activity and Bcl-2 expression leading to induction of radio sensitivity in human prostate cancer cells PC-3. Cancer biology & therapy 1(2): 152-160.
-
Ranganathan P, Rangnekar VM (2005) Regulation of cancer cell survival by Par-4. Ann N Y Acad Sci 1059: 76-85.
-
Goswami A, Burikhanov R, de Thonel A, Fujita N, Goswami M, et al. (2005) Binding and phosphorylation of par-4 by akt is essential for cancer cell survival. Mol cell 20(1): 33-44.
-
Moreno-Bueno G, Fernandez-Marcos PJ, Collado M, Tendero MJ, Rodriguez-Pinilla SM, et al. (2007) Inactivation of the candidate tumor suppressor par-4 in endometrial cancer. Cancer research 67(5): 1927- 1934.
-
Hebbar N, Shrestha-Bhattarai T, Rangnekar VM (2013) Par-4 prevents breast cancer recurrence. Breast cancer research : Breast Cancer Res 15(5): 314.
-
Shrestha-Bhattarai T, Hebbar N, Rangnekar VM (2013) Par(-4)oxysm in breast cancer. Cancer cell 24(1): 3-5.
-
Ahmed MM, Sheldon D, Fruitwala MA, Venkatasubbarao K, Lee EY, et al. (2008) Down regulation of PAR-4, a pro-apoptotic gene, in pancreatic tumors harboring K-ras mutation. International journal of cancer 122(1): 63-70.
-
Zhang L, Ren X, Cheng Y, Liu X, Allen JE, et al. (2014) The NF kappaB inhibitor, SN50, induces differentiation of glioma stem cells and suppresses their oncogenic phenotype. Cancer biol ther 15(5): 602-611.
-
Sharma AK, Sharma A, Desai D, Madhunapantula SV, Huh SJ, et al. (2008) Synthesis and anticancer activity comparison of phenylalkyl isoselenocyanates with corresponding naturally occurring and synthetic isothiocyanates. J Med Chem 51(24): 7820-7826.
-
Sharma AK, Kline CL, Berg A, Amin S, Irby RB, (2011) The Akt inhibitor ISC-4 activates prostate apoptosis response protein-4 and reduces colon tumor growth in a nude mouse model. Clinical cancer research: an official journal of the American Association for Cancer Research 17(13): 4474-4483.
-
Wischhusen J, Naumann U, Ohgaki H, Rastinejad F, Weller M (2003) CP-31398, a novel p53-stabilizing agent, induces p53-dependent and p53-independent glioma cell death. Oncogene 22(51): 8233-8245.
-
Welch WC, Morrison RS, Gross JL, Gollin SM, Kitson RB, et al. (1995) Morphologic, immunologic, biochemical, and cytogenetic characteristics of the human glioblastoma-derived cell line, SNB-19. In vitro cellular & developmental biology.-Animal 31(8): 610-616.
-
El-Guendy N, Zhao Y, Gurumurthy S, Burikhanov R, Rangnekar VM, et al. (2003) Identification of a unique core domain of par-4 sufficient for selective apoptosis induction in cancer cells. Mol cell biol 23(16): 5516- 5525.
-
Jagtap JC, Parveen D, Shah RD, Desai A, Bhosale D, et al. (2014) Secretory prostate apoptosis response (Par)-4 sensitizes multicellular spheroids (MCS) of glioblastoma multiforme cells to tamoxifen-induced cell death. FEBS open bio 5: 8-19.
- Cancer Diagnosis from RNA Sequence of Blood Cells by Using AI
- Field Cancerization in Oral Cavity, Case Report and Review of Literature. Oncologic Program Salud Integral Hospital, Managua, Nicaragua
- Identification of B Lymphocytes in Cancer Patient’s Blood
- A Case Report of a Breast Cancer Patient Developing Pneumonitis as a Result of Abemaciclib Therapy
- Immune Checkpoint Therapeutics for Today’s Fight and Beyond
- The Amalgamated Sophomore-Gonadoblastoma