A Comparative Approach Based on Molecular Docking Interaction of Citalopram and Tianeptine with Serotonin Transporter
The present study elucidated the relationship between the mechanism of action of selective serotonin reuptake inhibitors (SSRI, citalopram) and selective serotonin reuptake enhancer (SSRE, Tianeptine) by comparing their effects using molecular docking in silico. We demonstrated the binding of antidepressants to the crystal structure of the serotonin transporter protein (PDB ID; 6w2c). Molecular docking was done with the help of simultaneous multiple ligands docking using AutoDock Vina. The binding affinity of the compounds was determined by the length of the hydrogen bond, binding energy, and amino acid residue clusters. The whole chain of amino acids in the allosteric (S2) and substrate binding (S1) site cavities of SERT were analyzed using prankweb. Tianeptine was found to be the most effective ligand with the binding affinity Δ -9.0 kcal/mol > citalopram Δ -8.9 Kcal/mol > serotonin -6.8 kcal/mol. It is concluded that tianeptine exhibits strong binding affinity and stability to SERT compared to citalopram. Further, computational data strongly manifest the mode of action of the atypical antidepressant tianeptine that helps restore normal serotonergic neurotransmission.
Introduction
Chronic antidepressant treatment not only alter gene expression of the target receptors and transporters [1] but may also affect the whole central nervous system (CNS) by altering a wide range of physiological functions of the CNS [2, 3]. The complications ranging from dry mouth to very unpleasant (constipation, akathisia, sexual dysfunction), painful (acute dystonias), disfiguring (weight gain, tardive dyskinesia) and life‐threatening (myocarditis, agranulocytosis) are provoked in response to the use of broad-spectrum antipsychotic medications [4]. Besides the fundamental role of these medications research still lacks the pros and cons of even the most clinically prescribed antipsychotic drugs. Serotonin transporters (SERT) are the main target of serotonergic drugs to maintain synaptic serotonin leve0ls by controlling the transmission of ions and substrate across the neurons. SERT is part of the large family of Na/Cl dependent membrane transporters, containing 630
amino acid proteins and 12 transmembrane domains [5, 6]. In addition to its role in depression, it is also an important pharmaceutical target for the treatment of anxiety [7], autism [8], and obsessive-compulsive disorder [9].
SERT is a membrane-bound transporter with 12 transmembrane (TM) helices. The TM1 – 5 and TM6 -10 are the same having a pseudo-2-fold axis. TM1 and TM6 unwound in the middle which makes up the binding site along with TM3 and TM8, which are kinked around the binding site. 3D model of the human serotonin transporter was constructed via homology modeling to evaluate its binding affinity to serotonin [10] and via Internal Coordinate Mechanics (ICM) for testing eleven tryptamine derivatives including escitalopram [11] (Gabrielsen, Ravna et al. 2012). The crystal structure of SERT has been studied at high resolution with both centrally- bound and allosterically- bound ligands [12]. Decades of studies with leucine transporters have reported two substrate-binding sites in SERT. One is located at the orthosteric substrate binding site (S1) and the second allosteric site (named S2) is located in the extracellular vestibule for substrate binding [13, 14]. These two discrete binding sites exist in all three monoamine neurotransmitter transporters (MATs) (5HT, DAT, and NET) [15, 16]. The gating amino acid residues in the extracellular conformation of SERT exhibit orientation that differs from those in an outward-open conformation of human SERT bound to serotonin [17].
Selective serotonin reuptake inhibitor; citalopram contains escitalopram and R- citalopram in 1:1 ratio. Escitalopram has a ︣50 fold higher affinity for the human SERT compared to R-citalopram [18]. Previously, we have reported that acute citalopram and tianeptine (10mg/ kg) improve serotonin dysfunction by enhancing free tryptophan uptake from the periphery to the brain [19, 20]. Tianeptine is classified as a tricyclic antidepressant due to its heterocyclic nature. Two heteroatoms are incorporated i-e sulfur hence its name “tia” and nitrogen instead of carbon in the central ring and carry an aminoheptanoic side chain. The tricyclic nucleus has an electron donor heteroatom in position 5, and an electron acceptor atom in position 3 in the aromatic ring [21]. It is a drug of abuse and addiction and possesses antidepressant and anxiolytic characteristics in common with citalopram [22, 23]. It is also involved in the improvement of cognitive and neuroprotective effects on 5HT (5- hydroxytryptamine) mediated behavioral deficits and also acts on the somatic symptoms of depression particularly digestive system [24].
According to a molecular dynamics (MD) simulation study by Kortagere, et al. a four-point pharmacophore model was constructed and about three million small molecules were screened on this ligand-based model [25]. In addition, the ligand database of the National cancer institute was screened using another structure-based pharmacophore model based on six docking conformations of SSRI, and two compounds NSC175176 and NSC705841 were proposed to be potent [26]. Koldsø, et al. have observed changes in SERT conformations by using three different types of ligands; non-competitive inhibitor (ibogaine), competitive inhibitor (cocaine), and substrate (5HT). The authors reported that while cocaine preserves outward-facing conformation, 5HT and ibogaine change SERT conformation towards inward- facing at sampled time-scale [27]. Recent experiments on multiple computational methods showed that Ala169, Ala173, Thr439, Gly442 and Leu443 amino acid residues possess key interaction with fluoxetine, sertraline, 8PR, and 68P [28]. Davis et al. documented binding of paroxetine to SERT depends on the charge distribution in the cavity of SERT. Further, SERT- drug interaction was analyzed through molecular docking analysis on various synthetic compounds [29].
The monoaminergic theory is the keystone followed by the mechanism of action of selective serotonin reuptake blockers, however, tianeptine is atypical tricyclic antidepressant and its complex mechanism of action is still under investigation in the treatment and pathophysiology of depression. Therefore, the present study aims to elucidate the relationship between the mechanism of action of citalopram and tianeptine using molecular docking interaction to SERT that may uncover their potential targets in the treatment of depression.
Material and Methods
Multiple ligand simultaneous Docking
The multiple ligand simultaneous docking was performed that uses the empirical scoring function of Autodock vina. For multiple ligand simultaneous docking, two ligands were docked simultaneously. This empirical scoring function calculates the fitness of interaction by summing up the contribution of a number of individual turns, which is an important energetic factor in protein-ligand binding.
Preparation of Ligand
Tianeptine, a selective serotonin reuptake enhancer (Tianeptine Acid, Tianeptine n[INN];7-{(3-chloro-6- methyl-5,5- Dioxo- 11 H-Benzo [C]{2,1} Benzothiazepin -11- Y)Amino] Heptanoic Acid), citalopram (selective serotonin reuptake inhibitor) and 5HT were downloaded in SDF(2D) format from pub-Chem database. The structures of all drugs are shown in Figure 1.
Preparation of Macromolecule
The 3D crystal structure of the serotonin transporter (SERT) (PDB ID; 6W2C) was downloaded from the protein databank, RCSB (Research Collaboratory For Structural Bioinformatics) site in PDB Format (www.rcsb.org/pdb) bound to paroxetine analog in PDB format and saved.
Heteroatoms were removed from the protein crystal structure for the prevention of unwanted interaction while docking. In order to validate ligand pose with high binding affinity among various ligand poses of test ligands, x, y, and z attributes in the protein crystal structure of SERT transporter 6w2c were noted as shown in Figure 2.
Setting Docking Parameter
Molecular docking was performed using Autodock 4.0, Autodock Vina to predict maximum binding affinity between ligands and protein. The SDF (spatial data file) formats of the ligands were converted into PDBQT, Protein (P), Data Bank (DB) , partial charge (Q) , and Atom type (T) format using command prompt from open babel for further docking studies. The receptor was edited by removing water, ligand molecule, and heteroatoms like metal ions in autodock tools (ADT). Heteroatoms were removed from the protein crystal structure for the prevention of unwanted interaction while docking. Then the macromolecule was saved in PDBQT format after adding polar hydrogens and Kolman charges. The docking parameter file was generated with grid spacing 1 Å and dimensions of 34.526 X 175.662 X 165.11 Å with x= 60, y=60, and z=60 coordinates and saved as config_multi in a folder. The grids spacing used was set accordingly to keep enough space for ligands to be docked on the surface. Results were analyzed and a two-dimensional graphical depiction of best complexes was assessed using discovery studio visualization and the detailed analysis on the physicochemical properties including aminoacid interaction, hydrogen bonding, and hydrophobic interactions of the docked complexes were obtained by protein-ligand interaction profiler (PLIP), a fully automated protein-ligand interaction tool and lig Plot.
Prank Web can identify ligand binding site by determining sequence conservation and is available at http://prankweb. cz/ [30].
Results
Figure 3 shows docking poses generated by autodock Vina. Tianeptine in complex with SERT shows the highest binding energy -9kcal/mol compared to citalopram -8.9 and to its substrate serotonin -6.8kcal/mol.
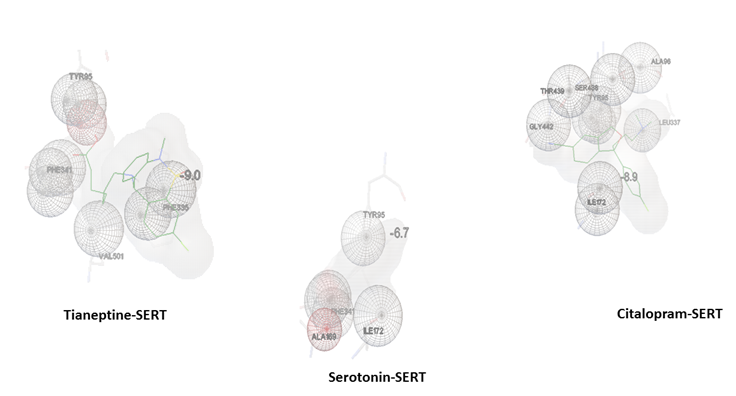
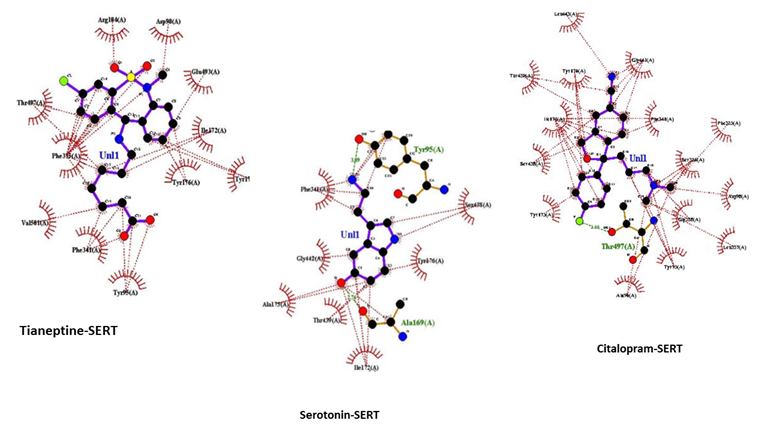
Figure 4 shows the amino acids involved in hydrophobic interaction of docked compounds obtained through ligplot+. The figure shows similarity in the amino acid residues involved in the hydrophobic interactions of citalopram and tianeptine to SERT are phe341, tyr95, tyr176, ile172, asp 98, phe335. Moreover, 5-HT-SERT complex shows hydrophobic interaction with amino acid residues similar to those of ligand (citalopram and tianeptine) complexes including ile172, tyr176, phe341. However, hydrophobic interaction shows some additional similarity within citalopram-SERT and serotonin-SERT complex by thr439 and gly442 amino acid residues.
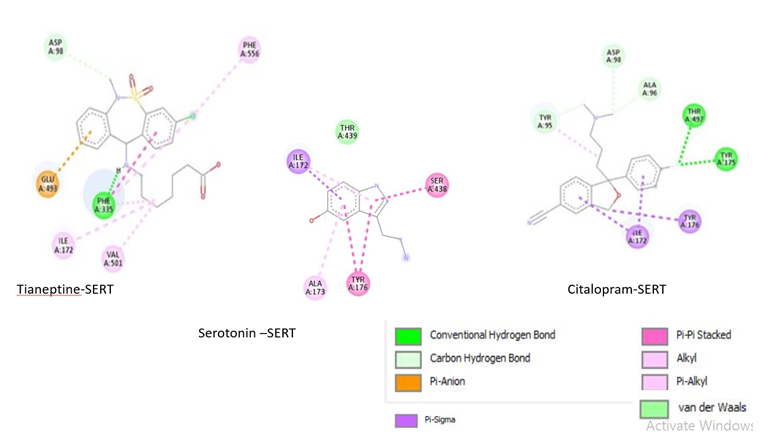
Figure 5: 2D interaction of SERT with ligands; Tianeptine-SERT. Citalopram-SERT .Serotonin –SERT obtained by discovery studio. Figure shows conventional hydrogen bond, Van der Waals forces, carbon hydrogen bond, Pianion, Pi-sigma, pi-pi- stacked, pi-alkyl bonds with lime green color, pale green color, light cyan color, yellow color, violet color, pink and light pink color respectively.
Figure 5 shows 2D representation of the docked complexes that determine steric forces involved in binding of ligands to SERT. Tianeptine-SERT and citalsopram-SERT complexes show alkyl, pi-alkyl, pi-anion and pi-sigma interaction respectively in addition to hydrogen and van der Waals forces. And, serotonin SERT complex shows alkyl, pi-alkyl, pi-sigma and pistacked bonds. Further, serotonin- SERT and citalopram-SERT complex have similarity in the amino acid residues tyr95, tyr176, and Ile172. However, Ile172 is the only common amino acid found in serotonin-, citalopram-, and tianeptine-SERT complex.
| Ligand Name | Binding affinity | No. of H bonds | Amino acid Residues with H bonding | Salt Bridges | Halogen Bonds | Hydrophobic interactions | π |
|---|---|---|---|---|---|---|---|
| ΔG (kcal/mol) | stacking | ||||||
| Tianeptine | -9 | …….. | …….. | 98ASP | 497THR | TYR95, ILE172, ILE172, ILE 172, TYR176, PHE 341 | ---- |
| Citalopram | -8.9 | 1 | THR-439 | 98ASP | 497THR | TYR95, ILE172, ILE172, ILE 172, TYR176, PHE 341 | ------ |
| Serotonin | -6.8 | 2 | 95-TYR, | --- | --- | ILE172, TYR175, TYR176, TYR176, PHE335, PHE341, PHE341, VAL501 | PHE335 |
| 335-PHE |
Table 1: Types of Secondary structures present in the 3D structure of SERT.
Table1: Physicochemical properties of hSERT and its inhibitors obtained by PLIP (protein-ligand interaction profile).
Table 1 shows physio-chemical properties including amino acid interaction, hydrogen bonding, and hydrophobic interactions of the docked complexes obtained through PLIP (protein-ligand interaction tool) fully automated protein- ligand interaction tool. None of the hydrogen bonds were seen with tianeptine –SERT, though citalopram-SERT complex showed one hydrogen bond with thr439. In addition, two hydrogen bonds 95tyr, 335phe were seen with serotonin- SERT interaction. Table shows name of amino acids involved in hydrophobic interactions with chain A of SERT. Both the ligands, citalopram and tianeptine, showed similar amino acid residues involved in hydrophobic interaction tyr95, ile172, tyr176, phe 341with chain A of SERT. Furthermore, SSRI and SSRE showed similarity in amino acid residues interaction with SERT involved in salt bridges 98asp and halogen bonds 497thr. However, serotonin showed hydrophobic interaction with some other amino acid residues like, tyr175, phe335 and val501.
| SR# | Type Of Secondary Structure | Percentage |
|---|---|---|
| 1 | Alpha Helix | 43.17% |
| 2 | 3 10 Helix | 0.00% |
| s03 | Pi Helix | 0.00% |
| 4 | Beta Bridge | 0.00% |
| 5 | Extended Strand | 19.85% |
| 6 | Beta Turn | 7.83% |
| 7 | Bend Region | 0.00% |
| 8 | Random Coil | 29.14% |
| 9 | Ambiguous States | 0.00% |
| 10 | Other States | 0.00% |
Table 2: Types of Secondary structures present in the 3D structure of SERT.
Table 2 shows percentage of forms in 3D structure as different forms of secondary structures later undergoes definite 3D form. Secondary structure prediction by SOPMA shows that sodium dependent serotonin transporter is composed of 549 amino acids and 43.17% alpha helix compared to beta turns (7.83%). There is no 310 helix, which is the most common type of secondary structure found in proteins and polypeptides as extensions of alpha helices found in either N or C terminal.
Table 3 shows the list of all amino acid residues obtained through Prankweb analysis that exist in the S1 (orthosteric) and ligand (S2/allosteric) binding sites. Results identified by prank web analysis are shown as a series of amino acid residues in the whole pocket of allosteric and orthosteric sites are represented as chains of 18 and 20 pockets respectively. There exist some difference between amino acid residues in serotonin and allosteric binding sites that are pocket 1; 152arg, 159lys, 167ileu, 237 leu, 453 glu, 454phe, 458trp, 468ala, 596arg, 599ile,600 thr, pocket2; 197 trp, 198 thr,210 thr,212 tyr, 213phe,220trp, 224ser,226ser, 227pro, 386met, 389met,390arg,391asn, 396glu,397val, 412glu,415ala, 416 asn, pocket 6; 184 leu,287phe, pocket 8 475phe, 577leu,584 ser, pocket 9 300ala pocket 10 157ile, 158phe, ,510val,515 phe, 593ile,597leu,604phe,608 ile,612ile, pocket14; (503thr, 583thr) 17251 ile, 253 trp,256ala, 257leu,478 leu, pocket 18 142tyr,143his,153lys,353phe, 516tyr,520gln, 524asp, 527glu, 614 pro , and 20 188ile, 254gln, 258 cys. However, many common amino acids are also listed in Table 3.
| Amino acid residues | Citalopram/tianeptine (allosteric S2) | Serotonin (orthosteric S1) |
|---|---|---|
| Pocket1 | 95 TYR,96 ALA,98ASP,99 LEU,100 GLY,103 TRP,104 ARG,108 ILE,166 CYS,169 ALA, 170 PHE,172ILE,173 ALA,175 TYR,176 TYR,179 ILE,182 TRP, 327 ILE,328ASP, 331ALA, 332 GLN, 335 PHE, 336 SER, 338 GLY,339 PRO,341PHE, 402GLY,403PRO, 406LEU, 407PHE,438 SER,439THR, 442GLY,443 LEU,446VAL,465PHE,469 VAL, 472THR,473CYS, 486ALA, 489VAL, 490LYS, 493GLU, 494 GLU, 497THR,498 GLY,499PRO, 501VAL, | 95 TYR ,96 ALA, 98 ASP, 99 LEU, 100 GLY, 103 TRP, 104 ARG, 108ILE,152ARG, 159LYS, 163TYR, 166 CYS, 167 ILE,169 ALA,170 PHE,172 ILE,173 ALA, 175 TYR, 176 TYR,179 ILE, 182 TRP,237 LEU,327 ILE, 328 ASP, 331 ALA, 332 GLN, 335 PHE, 336 SER, 338 GLY, 339 PRO, 341PHE,402 GLY,403PRO, 406 LEU,407 PHE, 438SER, 439 THR, 442GLY,443LEU,446VAL,450VAL,453 GLU,454PHE, 458TRP, |
| 556 PHE,559 SER,561PRO,565 LEU | 465PHE,468ALA,469 VAL,472THR,473CYS,486ALA, 489VAL,490 LYS,493GLU,494GLU,497THR,498GLY, 499PRO, 501 VAL, 556 PHE, | |
| - | 559 SER,561 PRO,565 LEU,566 PHE,588 CYS,591 THR,592 TYR,595 TYR,596 ARG,599 ILE,600 THR |
Table 3: List of all amino acid residues obtained through Prankweb analysis in the allosteric (S2) and orthosteric (S1) cavities.
197 TRP, 198 THR,210 THR,212 TYR, 213 PHE, 220TRP,
Pocket2 105 PHE,118LEU,121TYR122 THR,125 ALA, 126 ILE, 130 ILE, 366 VAL,369 CYS370, MET,373 PHE
- 105 PHE,118 LEU,121 TYR,122 THR,125 ALA,126
- ILE,130 ILE,362 LEU,366 VAL,369CYS,370 MET,373
- PHE
- Pocket3
- 123 ILE, 124MET127, PHE,323 THR,326 TRP, 544
- ILE,547 LEU, 548 PHE, 551PHE
- Pocket4
- 139 LEU,147CYS 150 ILE, 151TRP, 154 ILE,161 ILE
- 123 ILE, 124 MET,127PHE,326TRP,544 ILE,547 LEU,
- 548PHE,551PHE
- 165 ILE, 345 LEU, 508 GLU, 512 VAL
- 79ARG,80 GLU,272 LYS,
- 79 ARG,80GLU,272 LYS,274VAL,350TYR,448
- THR,451 LEU,452 ASP,459ALA,462ARG
- 274 VAL,350 TYR, 448THR,451 LEU, 452 ASP,459
- ALA
- Pocket 5
- 462ARG
- 184 LEU,187LEU,261 LEU,262ILE, 265VAL,
- Pocket6
- 186LEU,187 LEU,261LEU,262 ILE, 265VAL,280 VAL,
- 283VAL, 284 THR, 429 LEU,432 ILE,433THR, 436
- LEU
- 280 VAL, 283VAL,284 THR, 287 PHE,429, LEU,432
- ILE,433 THR,436 LEU
- Pocket 7
- 163 TYR,450 VAL
- 139LEU,147CYS,150 ILE,151 TRP,154 ILE,161
- ILE,165 ILE, 345 LEU,508GLU, 512VA
- 171TYR,174 SER,475 PHE,476 GLY,479VAL,480
- THR,488VAL,491 LEU,492LEU,577LEU,581 ILE,584
- SER
- Pocket 8
- 171TYR,174SER,476GLY,479VA,480 THR,
- 488VAL,491LEU,492 LEU,581 ILE
- Pocket 9
- 495TYR,557LEU, 560 PRO,579 TYR, 582 GLY
- 296 LEU,300ALA, 309VAL,313 LEU,374
- VAL,377PHE,381THR
- Pocket 10
- 164ALA,168 ILE,500ALA,504 VAL,507 ILE,586
- PHE,589 ILE
- 157ILE,158 PHE,507ILE,510VAL,515 PHE,593
- ILE,597 LEU,604 PHE,608 ILE,612ILE
- Pocket11
- 229 GLU,400 ASP, 408 ILE, 409THR,566 PHE,567
- GLN
- 358TYR,536PHE, 537 TRP
- Pocket12
- 296 LEU,309 VAL,313LEU, 374VAL, 377 PHE
- 260 MET,263 PHE ,264THR,470VAL,471ILE, 474PH
- Pocket13
- 535 TRP, 536 PHE, 539ILE
- 164ALA,168ILE,500ALA,504VAL,586 PHE,589ILE
- Pocket14
- 362 LEU,537 TRP
- 503 THR, 557 LEU, 579 TYR,582GLY, 583THR
- Pocket15
- 260 MET,263 PHE,264 THR, 470VAL,471 ILE,
- 535 TRP,539 ILE
- 474PHE
- Pocket16
- 588 CYS,591 THR,592 TYR,595 TYR
- 90 LEU,93ILE,285ALA,286 THR,289 TYR,367 VAL
- Pocket17
- 90LEU,93ILE, 285 ALA,286 THR, 289TYR,367 VAL
- 251 ILE,253 TRP,256 ALA, 257LEU,478 LEU
- 142TYR,143HIS,153LYS,353 PHE,516TYR,520
- GLN,524ASP,527 GLU,614 PRO
- Pocket18
- 134 TYR,358 TYR, 525VAL, 529 LEU, 531 PHE, 533
- PRO, 534 GLY
- Pocket 19
- 134 TYR,525VAL,529 LEU,531 PHE,533 PRO,534GLY
- Pocket 20
- 188ILE, 254GLN,258 CYS
Table 4: List of all amino acid residues obtained through Prankweb analysis in the allosteric (S2) and orthosteric (S1) cavities.
Discussion
Drug response is a result of chemical interaction between drug and its binding site. In relevance to the mechanism of action of citalopram to increase serotonin levels in the synaptic cleft, molecular docking data shows lower binding (-8.9kcal/mol) affinity (docking score) by the citalopram to SERT compared to tianeptine (-9.0 kcal/mol) with the lower root mean square deviation (RMSD) (0.00). It was reported that if the RMSD values of the best conformation is < 2.0Å from the bound ligand in the experimental crystal structure then the used scoring function is successful.
In contrast, previous studies reported earlier that the tianeptine exhibits a very low affinity for serotonin transporter and appears to not increase its mRNA expression, alter the firing rate of serotonergic neurons, and the activity of 5HT1A receptors in the dorsal raphe region. Therefore, it seems unlikely that it can increase serotonin reuptake [31] and works beyond the monoaminergic theory of depression [32, 33].
In view of the present finding, it can be suggested that the binding mode of tianeptine to SERT differentiates it from the mechanism of action of citalopram. In addition, none of the hydrogen bonds seemed to be formed by the tianeptine– SERT protein complex. However, citalopram-SERT showed one hydrogen bond (Thr439) (Table 1). It could be due to the reason that tianeptine lacks a net physiological charge (zero) compared to citalopram having a +1 physiological charge. A study on the mechanism of paroxetine inhibition to SERT suggested that the binding of paroxetine depends on the charge distribution of the cavity [29]. Hydrogen bonding in donor and acceptor groups creates minor changes in electron distribution that affects the shape and depth of the wells corresponding to covalent modification of the protein [34]. And, the chemical structure and moiety of both the drugs and receptors are important and a small change can result in a diminished response.
It has been suggested that the serotonergic system has not been implicated in the antidepressant mechanism of action of tianeptine [35]. However, numerous experimental and clinical studies have documented that the citalopram facilitates serotonergic neurotransmission [36] and its additional anxiolytic effects had been reported [20]. Hyttel, 1977 reported that citalopram decreases serotonin synthesis and turnover possibly by increasing its concentration in the synaptic cleft. This increase of concentration in the cleft, in turn, decreases 5-HT neuronal firing as a result of feedback inhibition. It was reported that the citalopram at doses that predominantly affect serotonin re-uptake, did not appear to alter the endogenous levels of serotonin, norepinephrine, and dopamine in the rat brain [37].
In contrast, previously demonstrated experiments on the uptake of serotonin in the rat brain synaptosomes differentiate the mechanism of action of tianeptine from selective serotonin reuptake inhibitors and that suggests its characteristic to reuptake synaptic serotonin [38]. Tianeptine appeared to reduce hypothalamic pituitary adrenal (HPA) axis activity and cortisol release when given several hours before restraint stress [39]. With these effects, tianeptine shows anxiolytic effects [40, 24]. Further, it was found earlier that chronic treatment with different classes of antidepressants including selective serotonin reuptake enhancer decreases the stress-induced level of plasma corticosterone [41, 42]. Some additional effects of tianeptine on neuroprotection have been observed that appear to restore neuronal plasticity [43, 44]. Chronic treatment of tianeptine manifests its sensitizing effects on alpha 1 adrenergic receptors [45, 46].
Our, present study suggest that the neutrality of tianeptine exhibits more affinity and binding stability for SERT compared to citalopram. Further, due to its unique structural characteristics, it may possess a broad binding affinity with multiple neuronal systems in addition to the serotonergic system. In addition, both the drugs show similarity in amino acid residues Asp 98, Thr 497 involve in halogen bond and salt bridge formation. Further, a 2D diagram of non-covalent and other relevant interactions shows steric hindrance that determine the affinity to which ligands interact with the SERT. Citalopram and serotonin interaction to SERT shows similarity in the amino acid residues tyr95, tyr176, and Ile172. And, Ile172 is the only common amino acid found in all three complexes. In relation to both the ligands, the serotonin-SERT complex shows the lowest binding affinity (-6.0 kcal/mol) and two hydrogen bonds (Tyr95, Phe335).
In addition to drug specificity, interaction to ligands also depends on the primary amino acid sequence and the stability of the structure of receptor. Since the beta form is more stable compared to alpha form, the percentage of alpha helix was found to be 43% compared to beta turn (7%) in SERT (Table 2). Therefore, by looking into the percentage of forms in secondary structure prediction that later undergoes 3D structure of SERT, it can be suggested that the 3D structure of SERT is not stable. And, this structural instability favours the binding of sodium dependent serotonin transporter to the ligands non-covalently along with other steric forces to elicit pharmacological effect produced by the drug. We are the first to report the chain of amino acids in orthosteric substrate-binding S1 and allosteric S2 sites through pranweb analysis (Table 3). We found similarities in the chains of the amino acids present in the pockets of cavities formed by either inhibitor (citalopram/tianeptine). However, some non-polar and neutral amino acids 323thr (pocket 3), 186leu
(pocket 6), 495tyr, and 560pro (pocket 6) were found to be crucial to the allosteric binding site. Gln 332 was found to be one of the most important residues around the entrance of SERT analyzed by the cysteine accessibility method [47]. With these reports, Gln 332 residue appears to be found in the entrance (pocket 1) of both substrate and allosteric binding sites. It can be suggested that the Gln being its polar nature may participate in the binding of ligand and receptor and therefore the amino acid residues with their chiral nature and the charge on them encounter specific cavities in the SERT that allows binding inhibitors or substrate.
Previous findings through mutagenesis and molecular modeling reported that Lys99, Arg104, Trp103, Ile179, Ala486, Val489, Lys490, and Gly402 predominantly reside around the allosteric binding site [48]. Similarly, Prank web analysis in our present results shows the detailed location of these residues (Leu 99, Arg104, Trp103, Ile179, Gly402, Ala486, Val489, Lys490) in the allosteric binding site, which appears to exist in pocket 1. It can be suggested that these residues located in the allosteric region are specific to the entrance of ligand molecules.
Our present results confirm the presence of Tyr176, Phe335 and Ser336 resided in the pocket of the substrate- binding site as represented in Table 2. In line with these findings, previous investigations reported that amino acid residues Tyr176 and Phe335 and Ser 336 are important amino acid residues for 5HT binding in the transmembrane domain 6 (TM6) of hSERT [49, 50]. Amino acids that are involved in the hydrophobic interactions by both tianeptine and citalopram show similarity i-e Tyr95, Ile172, Phe 341, Tyr 176 (Table 2, Figure 6). Similar to our findings, Henry et al. (2006) identified Tyr95 and Ile172 amino acid residues as major determinants of binding for several antagonists in the human serotonin transporter [51]. Our findings suggest that these amino acids are crucial to the interaction of serotonergic drugs with SERT. Similarly, mutational and computational efforts found Asp98, Asn177, Phe341, and Ser438 amino acid residues were crucial in the binding pocket [52].
Conclusion
It is concluded that the tianeptine exhibits strong binding affinity and stability to SERT compared to citalopram. Further, computational data strongly justifies the mode of action of the atypical antidepressant tianeptine to increase serotonin reuptake that helps restore normal serotonergic neurotransmission.
Contributions
IA and GA performed conceptual design of the study, IA and SB preformed and analyzed experiments, IA and GA prepared the manuscript. IA performed performed docking,. All authors have read and approved the final version of the manuscript.
References
-
Detke MJ, Johnson J, Lucki I (1997) Acute and chronic antidepressant drug treatment in the rat forced swimming test model of depression. Exp Clin Psychopharmacol 5(2): 107-112.
-
Lesch KP, Heils A (2000) Serotonergic gene transcriptional control regions: targets for antidepressant drug development? Int J Neuropsychopharmacol 3(1): 67-79.
-
Alex KD, Pehek EA (2007) Pharmacologic mechanisms of serotonergic regulation of dopamine neurotransmission. Pharmacol Ther 113(2): 296-320.
-
Stroup TS, Gray N (2018) Management of common adverse effects of antipsychotic medications. World Psychiatry 17(3): 341-356.
-
Ravna AW, Jaronczyk M, Sylte I (2006) A homology model of SERT based on the LeuT(Aa) template. Bioorg Med Chem Lett 16(21): 5594-5597.
-
Nencetti S, Mazzoni MR, Ortore G, Lapucci A, Giuntini J, et al. (2011) Synthesis, molecular docking and binding studies of selective serotonin transporter inhibitors. Eur J Med Chem 46(3): 825-834.
-
Murphy DL, Lerner A, Rudnick G, Lesch KP (2004) Serotonin transporter: gene, genetic disorders, and pharmacogenetics. Mol Interv 4(2): 109-123.
-
Sutcliffe JS, Delahanty RJ, Prasad HC, McCauley JL, Han Q, et al. (2005) Allelic heterogeneity at the serotonin transporter locus (SLC6A4) confers susceptibility to autism and rigid-compulsive behaviors. Am J Hum Genet 77(2): 265-279.
-
Delorme R, Betancur C, Wagner M, Krebs MO, Gorwood P, et al. (2005) Support for the association between the rare functional variant I425V of the serotonin transporter gene and susceptibility to obsessive compulsive disorder. Mol Psychiatry 10(12): 1059-1061.
-
Celik L, Sinning S, Severinsen K, Hansen CG, Moller MS, Bols M, et al. (2008) Binding of serotonin to the human serotonin transporter. Molecular modeling and experimental validation. J Am Chem Soc 130(12): 3853- 3865.
-
Gabrielsen M, Ravna AW, Kristiansen K, Sylte I (2012) Substrate binding and translocation of the serotonin transporter studied by docking and molecular dynamics simulations. J Mol Model 18(3): 1073-1085.
-
Coleman JA, Green EM, Gouaux E (2016) X-ray structures and mechanism of the human serotonin transporter. Nature 532(7599): 334-339
-
Zhao Y, Terry DS, Shi L, Quick M, Weinstein H, et al. (2011) Substrate-modulated gating dynamics in a Na+- coupled neurotransmitter transporter homologue. Nature 474(7349): 109-113.
-
Larsen MA, Plenge P, Andersen J, Eildal JN, Kristensen AS, et al. (2016) Structure-activity relationship studies of citalopram derivatives: examining substituents conferring selectivity for the allosteric site in the 5-HT transporter. Br J Pharmacol 173(5): 925-936.
-
Kristensen AS, Andersen J, Jorgensen TN, Sorensen L, Eriksen J, et al. (2011) SLC6 neurotransmitter transporters: structure, function, and regulation. Pharmacol Rev 63(3): 585-640.
-
Manepalli S, Surratt CK, Madura JD, Nolan TL (2012) Monoamine transporter structure, function, dynamics, and drug discovery: a computational perspective. AAPS J 14(4): 820-831.
-
Hellsberg E, Ecker GF, Stary-Weinzinger A, Forrest LR (2019) A structural model of the human serotonin transporter in an outward-occluded state. PLoS One 14(6): e0217377.
-
Sanchez C (2006) The pharmacology of citalopram enantiomers: the antagonism by R-citalopram on the effect of S-citalopram. Basic Clin Pharmacol Toxicol 99(2): 91-95.
-
Bano S, Gitay M, Ara I, Badawy A (2010) Acute effects of serotonergic antidepressants on tryptophan metabolism and corticosterone levels in rats. Pak J Pharm Sci 23(3): 266-272.
-
Ara I, Bano S (2012) Citalopram decreases tryptophan 2,3-dioxygenase activity and brain 5-HT turnover in swim stressed rats. Pharmacol Rep 64(3): 558-566.
-
Stephen MS (2017) Prescriber’s guide: Antidepressants: Stahl’s essential psychopharmacology. Cambridge University Press.
-
Hindmarch I (2001) Expanding the horizons of depression: beyond the monoamine hypothesis. Hum Psychopharmacol 16(3): 203-218.
-
Iffat Ara Amjad, SBAR (2019) Citalopram and Tianeptine: Pharmacological Interventions in alcohol withdrawal-induced negative mood states and serotonin insufficiency in Albino Wistar male rats. Indonesian J Pharm 30(2): 75-83.
-
Bailey SJ, Almatroudi, A., Kouris (2017) A. Tianeptine: An atypical antidepressant with multimodal pharmacology. Current Psychopharmacology 6(2): 94-110.
-
Kortagere S, Fontana AC, Rose DR, Mortensen OV (2013) Identification of an allosteric modulator of the serotonin transporter with novel mechanism of action. Neuropharmacology 72: 282-290.
-
Zhou ZL, Liu HL, Wu JW, Tsao CW, Chen WH, et al. (2013) Combining structure-based pharmacophore and in silico approaches to discover novel selective serotonin reuptake inhibitors. Chem Biol Drug Des 82(6): 705-717
-
Koldso H, Autzen HE, Grouleff J, Schiott B (2013) Ligand induced conformational changes of the human serotonin transporter revealed by molecular dynamics simulations. PLoS One 8(6): e63635.
-
Xue W, Wang P, Li B, Li Y, Xu X, et al. (2016) Identification of the inhibitory mechanism of FDA approved selective serotonin reuptake inhibitors: an insight from molecular dynamics simulation study. Phys Chem Chem Phys 18(4): 3260-3271.
-
Davis BA, Nagarajan A, Forrest LR, Singh SK (2016) Mechanism of Paroxetine (Paxil) Inhibition of the Serotonin Transporter. Sci Rep 6:23789.
-
Jendele L, Krivak R, Skoda P, Novotny P, Hoksza (2019) PrankWeb: a web server for ligand binding site prediction and visualization. Nucleic Acids Res 47(W1): W345-W349.
-
Malagie I, Deslandes A, Gardier AM (2000) Effects of acute and chronic tianeptine administration on serotonin outflow in rats: comparison with paroxetine by using in vivo microdialysis. Eur J Pharmacol 403(1-2): 55-65.
-
Kato G, Weitsch AF (1988) Neurochemical profile of tianeptine, a new antidepressant drug. Clin Neuropharmacol 11 Suppl 2: 43-50.
-
Svenningsson P, Tzavara ET, Witkin JM, Fienberg AA, Nomikos GG, et al. (2002) Involvement of striatal and extrastriatal DARPP-32 in biochemical and behavioral effects of fluoxetine (Prozac). Proc Natl Acad Sci U S A 99(5): 3182-3187
-
Orlowski R, Clark JA, Derr JB, Espinoza EM, Mayther MF, et al. (2021) Role of intramolecular hydrogen bonds in promoting electron flow through amino acid and oligopeptide conjugates. Proc Natl Acad Sci USA 118(11): e2026462118
-
McEwen BS, Chattarji S, Diamond DM, Jay TM, Reagan LP, et al. (2010) The neurobiological properties of tianeptine (Stablon): from monoamine hypothesis to glutamatergic modulation. Mol Psychiatry 15(3): 237-249.
-
Inoue T, Tsuchiya K, Koyama T (1996) Serotonergic activation reduces defensive freezing in the conditioned fear paradigm. Pharmacol Biochem Behav 53(4): 825- 831.
-
Hyttel J (1977) Neurochemical characterization of a new potent and selective serotonin uptake inhibitor: Lu 10- 171. Psychopharmacology (Berl) 51(3): 225-233.
-
Fattaccini CM, Bolanos-Jimenez F, Gozlan H, Hamon M (1990) Tianeptine stimulates uptake of 5-hydroxytryptamine in vivo in the rat brain. Neuropharmacology 29(1): 1-8.
-
Delbende C, Contesse V, Mocaer E, Kamoun A, Vaudry H (1991) The novel antidepressant, tianeptine, reduces stress-evoked stimulation of the hypothalamo-pituitary- adrenal axis. Eur J Pharmacol 202(3): 391-396.
-
Wagstaff AJ, Ormrod D, Spencer CM (2001) Tianeptine: a review of its use in depressive disorders. CNS Drugs 15(3): 231-259.
-
Reul JM, Labeur MS, Grigoriadis DE, De Souza EB, Holsboer F (1994) Hypothalamic-pituitary-adrenocortical axis changes in the rat after long-term treatment with the reversible monoamine oxidase-A inhibitor moclobemide. Neuroendocrinology 60(5): 509-519.
-
Reul JM, Stec I, Soder M, Holsboer F (1993) Chronic treatment of rats with the antidepressant amitriptyline attenuates the activity of the hypothalamic-pituitary- adrenocortical system. Endocrinology 133(1): 312-320.
-
Gomez E, Catalan R, Navines R, Gasto C (2001) Serotonin receptor changes in depression: evidences and limitations. Actas Esp Psiquiatr 29(3): 186-194.
-
McEwen BS, Chattarji S (2004) Molecular mechanisms of neuroplasticity and pharmacological implications: the example of tianeptine. Eur Neuropsychopharmacol 14 Suppl 5: S497-502.
-
Rogoz Z, Skuza G, Dlaboga D, Maj J, Dziedzicka- Wasylewska M (2001) Effect of repeated treatment with tianeptine and fluoxetine on the central alpha(1)- adrenergic system. Neuropharmacology 1(3): 360-368
-
Alamo C, Garcia-Garcia P, Lopez-Munoz F, Zaragoza C (2019) Tianeptine an atypical pharmacological approach to depression. Rev Psiquiatr Salud Ment Engl Ed 12(3): 170-186.
-
47.Torres-Altoro MI, Kuntz CP, Nichols DE, Barker EL (2010) Structural analysis of the extracellular entrance to the serotonin transporter permeation pathway. J Biol Chem 285(20): 15369-15379.
-
Plenge P, Shi L, Beuming T, Te J, Newman AH, et al. (2012) Steric hindrance mutagenesis in the conserved extracellular vestibule impedes allosteric binding of antidepressants to the serotonin transporter. J Biol Chem 287(47): 39316-39326.
-
Chen JG, Sachpatzidis A, Rudnick G (1997) The third transmembrane domain of the serotonin transporter contains residues associated with substrate and cocaine binding. J Biol Chem 272(45): 28321-28327.
-
Sinning S, Musgaard M, Jensen M, Severinsen K, Celik L, et al. (2010) inding and orientation of tricyclic antidepressants within the central substrate site of the human serotonin transporter. J Biol Chem 285(11): 8363-8374.
-
Henry LK, Field JR, Adkins EM, Parnas ML, Vaughan RA, et al. (2006) Tyr-95 and Ile-172 in transmembrane segments 1 and 3 of human serotonin transporters interact to establish high affinity recognition of antidepressants. J Biol Chem 281(4): 2012-2023.
-
Andersen J, Olsen L, Hansen KB, Taboureau O, Jorgesnsen FS, et al. (2010) Mutational mapping and modeling of the binding site for (S)-citalopram in the human serotonin transporter. J Biol Chem 285(3): 2051-2063.
- Acido Labile or Gastro Irritant Apis and Enteric Release in Galenic Practice: An Overview
- A Study on Knowledge, Attitude and Practice of Hand Hygiene among Healthcare Professionals at a Tertiary Care Hospital, India
- Influence of Inoculum Concentration on In Vivo Incubation Period of Emmia lacerata, Pathogenesis and Management of Wilt in Pepper (Capsicum annuum L.)
- Vanilla’s Chemistry
- Marine Anti-Cancer Compounds and Adverse Effects of Global Warming on Oceans: An Overview
- Serological Investigation of Chikungunya Virus Antibody among Malaria-Suspected Febrile Patients in Some Healthcare Facilities in Rivers State