Hematopoietic Stem Progenitor Cells Reduce the Proliferation of Tregs via Regulation of IL-12/STAT4 Signaling During Chronic Stress
Physical or psychological chronic stress mediates the impaired immune response by inducing the proliferation of regulatory T cells (Tregs). Our recent studies showed that hematopoietic stem progenitor cells (HSPCs) significantly modulated the immune system. HSPCs prevented stress-induced lymphocyte apoptosis, exerting a protective effect in chronic stress-induced immune responses. Thus, whether HSPCs could prevent the proliferation of Tregs in conditions of chronic stress was assessed in the present study. Injection of HSPCs inhibited the proliferation of Tregs and reduced the expression of inhibitory molecules of Tregs under conditions of chronic stress. Additionally, the protective effects of HSPCs were shown to be dependent on IL12. IL-12/STAT4 signaling in the modulation of proliferation of Tregs induced by chronic stress. When HSPC injection was co-administered with an IL-12 neutralizing antibody or selective STAT4 inhibitor in chronic stressed mice, the protective effects of HSPCs were abrogated. Injection of HSPCs increased IL-12 production by splenocytes in chronic stressed mice. These results suggest that HSPCs inhibit the proliferation of Tregs and this is dependent on IL-12 production to protect the impaired immune response.
Introduction
Epidemiological studies have suggested that physical and psychological stress leads to the impairment of the immune system and thus enhances susceptibility to diseases [1, 2]. The mechanisms include altered expression profiles of cytokines [3, 4], increased apoptosis of T cells and dendritic cells [4, 5], and enhanced proliferation of myeloid- derived suppressor cells [6]. It has been extensively shown that chronic stress induces the proliferation of regulatory T cells (Treg) [4, 7], which is at least in part due to immunosuppression.
CD34 is a marker of hematopoietic stem progenitor cells (HSPCs) [8]. Injection of HSPCs improved survival in late sepsis, and corrected immunosuppression through modulation of an impaired immune responses in late sepsis [9]. The beneficial effects of HSPCs may be related to their differentiated capacity compared with mature immunocompetent cells, as well as their ability to nest to the site of injury and inflammation [9]. Our previous study showed that HSPCs prevent chronic stress-induced immune impairment through modulation of a T cell-mediated immune response [10].
Tregs exert their suppressive function through the expression of inhibitory molecules such as cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), glucocorticoid- induced tumor necrosis factor (GTIR) and lymphocyte- activation gene 3, which decrease the proliferation of T cells by inhibiting the function of co-stimulatory markers [11].
Chronic restraint stress may increase the frequency and suppressive function of Tregs resulting in mediation of immunosuppression. Injection of HPSCs may thus offer a novel strategy to protect against a deleterious immune response induced by conditions of chronic stress. Therefore, it was speculated that HSPCs could prevent the proliferation of Tregs in chronic restraint stress to alleviate immune impairment.
Materials and Methods
Animals
BALB/c male mice were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. (China). Renmin Hospital of Wuhan University Animal Care and Use Committee approved all experimental procedures, which were performed in accordance with institutional animal welfare guidelines.
Establishment of the Chronic Restraint Stress Model
Male BALB/C mice, 6-7 weeks old, were used to establish the chronic physical restraint stress model previously used [4, 10, 12]. According to the protocol, mice were held in a 50-ml tapered centrifuge tube with several punctures for ventilation. Mice were kept horizontally in the tubes for 12 h followed by a 12-h rest. During the rest period food and water were provided. The mice in the control group were placed in their original cage providing food and water to the same schedule as for the stressed mice group. Mice were restrained for three cycles. After finishing restraint, mice were anesthetized by inhalation of 2.5% isoflurane (Abbott Pharmaceutical Co. Ltd.). Spleens were harvested. Blood and serum samples were collected for measurement of cytokine levels.
Isolation and Injection of CD34+ HSPCs in Mice
The bone marrow cells were flushed from the femurs and tibias using FBS-free RPMI 1640 medium under sterile conditions as described previously [9, 10]. The cells were repeatedly aspirated using a pipette to establish a single cell suspension and filtered using a 70-μm nylon strainer. The cells were aspirated again and filtered after erythrocyte lysis. The CD34+ HSPCs were separated using magnetic-assisted cell sorting [9, 10]. The single cell suspension was incubated with biotinylated mouse anti-CD34 antibody (eBioscience) for 20 min at 4˚C. The cells were incubated with antibiotin magnetic beads for 20 min at 4˚C after washing and then positive selection was applied using a Miltenyi selection column.
The isolated HSPCs were resuspended in sterile PBS, and mice were injected with HSPCs (1x106 in 100 μl PBS, >90% purity) or vehicle (heat-killed 1x106 HSPCs in 100 μl PBS) one day before the restraint. The control group was injected with PBS.
Reagents
Recombinant mouse (r)IL-12, rIL-4, rIL-7, TGF-β1 and a neutralizing antibody against mouse IL-12 were purchased from R&D systems. Goat polyclonal anti-mouse IL-12 IgG (0.01 mg/mouse) was administered by i.p. or the same amount of non-immune IgG as described previously [13, 14]. Either rIL-12(0.01mg/mouse) or neutralizing antibody against IL-12 (IL-12-Ab) was administrated before HSPC treatment. Five mice in the stressed group according to the random principle received daily i.p. injections of Lisofylline (LSF, 50 mg/kg, Enzo Life Sciences) [15] before the injection of HSPCs.
Isolation of CD4+ T cells and flow cytometry (FCM)
Single splenocyte suspensions were made after 3 cycles of 12-h restraint stress. CD4+ T cells were purified using a mouse CD4+ T cell kit (R&D Systems, Inc.) as described previously [4, 10]. CD25+ FOXP3+ cells from the purified CD4+ T cells were identified as Tregs. CD25+ FOXP3+ were analyzed by FCM as described in our previous study [4], labeled with the appropriate fluorochrome-conjugated antibodies. The samples were analyzed by FCM. Single splenocyte suspensions from unstressed or stressed mice were also stained with an anti-IL-12 antibody conjugated to APC (Biolegend, 1:50). The following antibodies were used: Anti-mouse-FOXP3 conjugated to phycoerythrin (PE) (Biolegend, 1:50), anti-mouse CD25 conjugated to fluorescein isothiocyanate (FITC) (Biolegend, 1:50).
CD4+CD25+ Treg sorting
Single splenocyte suspensions were prepared as described above. CD4+CD25+ regulatory T cells were isolated using a Regulatory T Cell Isolation Kit mouse (MACS, Miltenyi Biotec GmbH) according to the manufacturer’s instructions and as described previously [16]. First CD4+ T cells were isolated as described above, then they were labeled with an anti-CD25 antibody conjugated to Microbeads for positive selection.
Cell Culture
Cells (splenocytes and isolated HSPCs) were cultured in complete RPMI 1640 medium with 10% FBS, 2 mM l-glutamine, and 1x penicillin-streptomycin in a humidified incubator at 37˚C and supplied with 5% CO2 air. The cultured splenocytes were stimulated with rIL-12 (10ng/ ml), rIL-4 (10ng/ml), or rIL-7 (10ug/ml) and with different concentrations of TGF-β1 (10ng/ml, 20ng/ml and 30ng/ ml) for 24 h before evaluation of Tregs by FCM. The purified CD4+CD25+ T cells were cultured to detect the inhibitor molecules released from the stressed mice with or without HSPCs treatment. HSPCs were cultured with or without serum from the stressed or unstressed mice for the IL-12 assay.
Western Blot Analysis
Western blotting was performed as described previously [10]. Nuclear and cytoplasmic protein from spleens were extracted using a NE-PER Nuclear and Cytoplasmic Extraction Reagents (Thermo Fisher Scientific, Inc.). The membranes containing the resolved proteins were incubated with primary antibodies overnight at 4˚C. The primary antibodies used were bought from Cell signaling, including anti-phospho-STAT4 (1:500), anti-STAT4 (1:1,000), anti- FOXP3 (1:500), anti-GAPDH (1:1000), and anti-IL-12Rβ1 (R&D system,1:500).
ELISA. IL-12 in the serum were detected using mouse ELISA kits (R&D systems, Inc.).
Reverse transcription-quantitative PCR (RTq- PCR)
RNA was extracted using a Qiagen RNeasy kit (Qiagen
GmbH). RT-qPCR was used to determine the mRNA expression levels. qPCR was performed using SYBR Green Fluorescein PCR MasterMix (SABiosciences). GAPDH was used as the internal control. The sequences of the primers used were: CTLA-4 forward, 5’-CCG TGC CCA GAT TCT GAC-3’ and reverse, 5’-AAA CAA CCC CGA ACT AAC TGC-3’; GITR forward, 5’-GACCGAAGACGCCAGAAG-3’ and reverse, 5’-CTCACACCCACAGGTCTCT-3’;IL-12 forward, 5′-CAG AAG CTA ACC ATC TCC TGG TTTG-3′ and reverse, 5′-TCC GGA GTA ATT TGG TGC TTC ACA C-3′;GAPDH forward,5′-TGA CCA CAG TCC ATG CCA TC-3′ and reverse, 5′-GAT GGG GGT TAC ACA GGC AG-3′.
Statistical Analysis
All values are presented as the mean ± SD. Data were analyzed using GraphPad Prism 6 (GraphPad Software, Inc.). Significant two-way ANOVA results were further analyzed by a Bonferroni’s multiple comparison tests. P<0.05 was considered to indicate a statistically significant difference.
Results
HSPCs prevent the proliferation of Tregs and decrease the suppressive activity in chronic restraint stress. Injection of CD34+HSPCs before the initiation of restraint stress reduced the increase in the frequency of Tregs significantly (Figure 1A). Meanwhile, HSPC treatment reversed the stress-induced upregulation of FOXP3, which is a specific marker of Tregs (Figure 1B), suggesting that HSPCs prevented the proliferation of Tregs induced by chronic stress. CTLA-4 and GTIR on the surface of Tregs are responsible for the suppressive activity as they influence the stimulatory capacity of antigen presenting cells (APCs) [16]. Their expression is upregulated and is related to the suppressive activity of HSPCs after activation. Thus, the expression of CTLA-4 and GTIR in CD4+CD25+ Tregs sorted from stressed and unstressed mice with or without HSPC treatment was determined. The mRNA expression levels of CTLA-4 and GTIR were upregulated significantly in chronic stress (Figure 1C). Importantly, the percentage of CTLA-4 and GTIR separately stained with FOXP3 in sorted CD4+CD25+ Tregs increased in stressed mice, and HSPCs alleviated the upregulation in CTLA-4 and GTIR expression. These data suggest that HSPCs alleviated the suppressive activity of the CD4+CD25+Tregs.
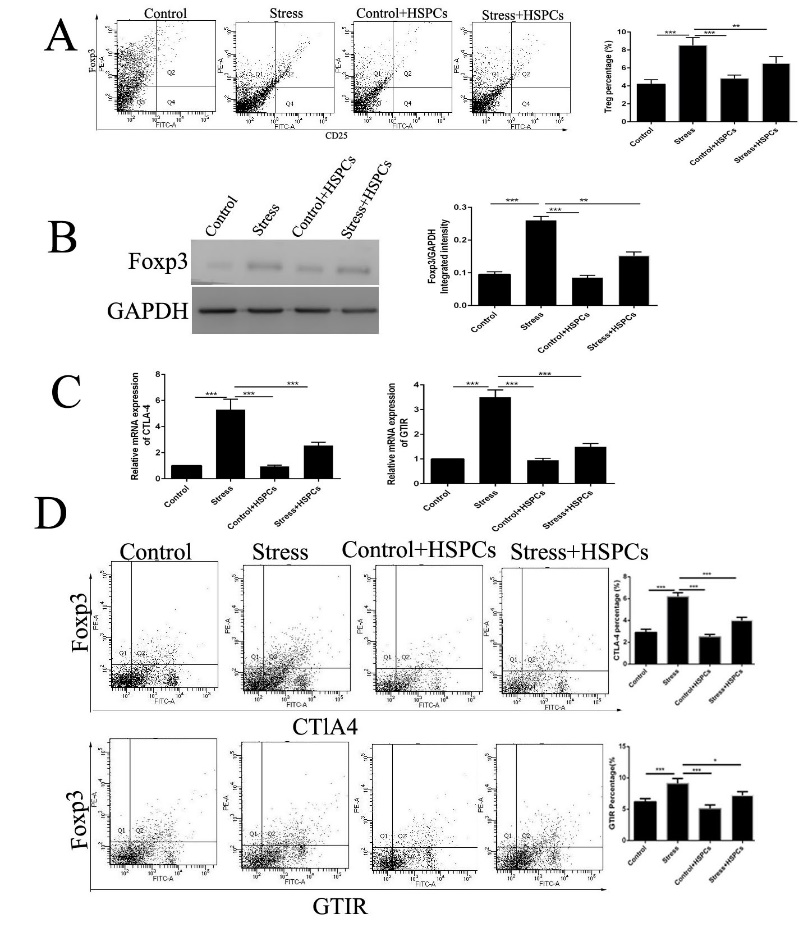
Figure 1: HSPCs reduce the proliferation and decrease the suppressive activity of Tregs in conditions of chronic stress. BALB/c male mice (6-8 weeks old) were injected intravenously with CD34+HSPCs (1x106) in 100 µl PBS or vehicle before the initiation of restraint stress. The mice were subjected to chronic restraint stress for 3 days, 12 h per day. (A) CD4+ cells in the spleens were isolated, and then stained with PE-FOXP3 and FITC-CD25. They were confirmed to be Tregs using flow cytometry. (B) FOXP3 expression in spleens was detected by western blotting. (C) CTLA-4 and GTIR in CD4+CD25+ cells were evaluated. D: The percentage of doubled stained CTLA-4 or GTIR and FOXP3 in sorted CD4+CD25+ regulatory T cells was determined. n =5 per group. P<0.01, *P<0.001. HSPC, hematopoietic stem progenitor cell; Tregs, regulatory T cells; FOXP3, forkhead box P3.
IL-12 abrogates the proliferation and suppressive activity of Tregs
IL-12 abrogates the increase in the number of Tregs in splenocytes stimulated with TGF-β1. To investigate which cytokines may mediate the protective effects of HPSCs, the expression of cytokines that facilitate or prevent the proliferation of Tregs were determined. Splenocytes were cultured with rIL-4, rIL-12, or rIL-7 for 24 h with different concentrations of TGF-β1 and the proportion of Tregs was determined using FCM. IL-12 was highly effective in reducing the frequency of Tregs among the tested cytokines, whereas other candidate cytokines showed minimal or no protective effect (Figure 2A).
rIL-12 Reduced the Frequency of Tregs invivo
Previous studies reported that IL-12 stimulated IFN- γ-mediated inhibition of tumor-induced proliferation of regulatory T-cells [17]. To further evaluate whether IL-12 was responsible for the prevention of proliferation Tregs in chronic restraint stress, rIL-12 was injected before the initiation of stress, and it abrogated the proliferation of Tregs in vivo (Figure 2B). IL-12 also reduced the expression of inhibitory molecules in Tregs (Figure 2C). Furthermore, rIL-12 treatment downregulated FOXP3 expression and increased the expression of p-STAT4 in spleens (Figure 2D), consistent with reports that IL-12 induces Tregs to produce IFN-γ and this is dependent on STAT4 signaling [17, 18].
Taken together, these studies demonstrate that IL-12 inhibits the activation of Tregs.
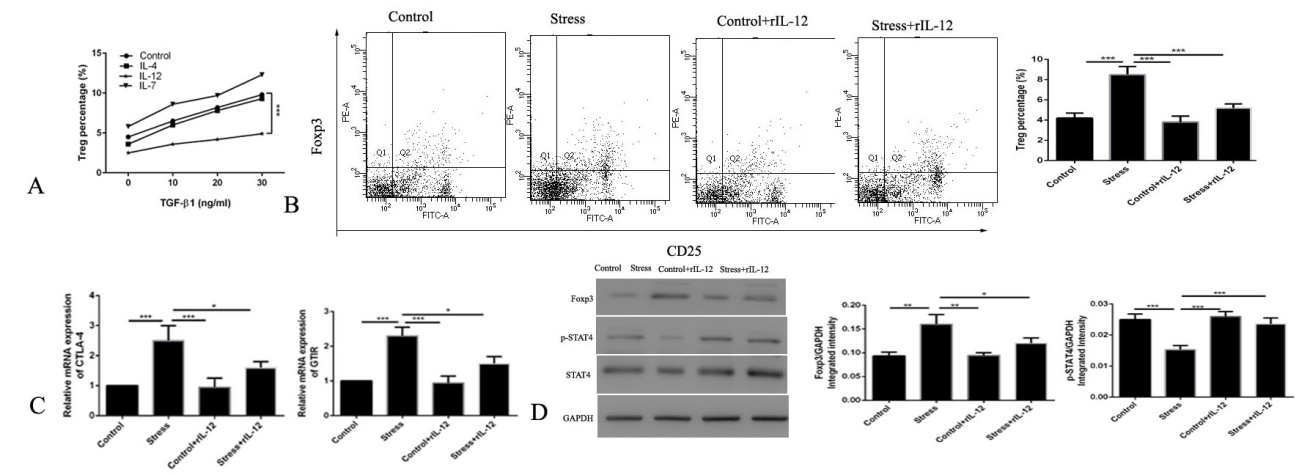
Figure 2: IL-12 exerts potent protection against chronic restraint stress-induced proliferation of Tregs. (A) The proportion of Tregs were determined using flow cytometry in the splenocytes after culturing with different concentrations of TGF-β1 and treatment with different cytokines (10 ng/ml rIL-12, rIL-4, or rIL-7). rIL-12 blocked the (B) proliferation of Tregs and (C) expression of inhibitory molecules in chronic stressed mice. (D) The expression of FOXP3 and p-STAT4 were examined by western blotting with or without rIL-12 treatment before chronic stress. n=5 per group. ***P<0.001. r, recombinant; Tregs, regulatory T cells; FOXP3, forkhead box P3.
The Protective Effect of HSPCs is Dependent on IL-12/STAT4 Signaling
To evaluate whether IL-12/STAT4 signaling is critical in the protective effects of HPSCs in chronic stress, IL-12-Ab or selective STAT4 inhibitor LSF were administrated before the injection of HSPCs.
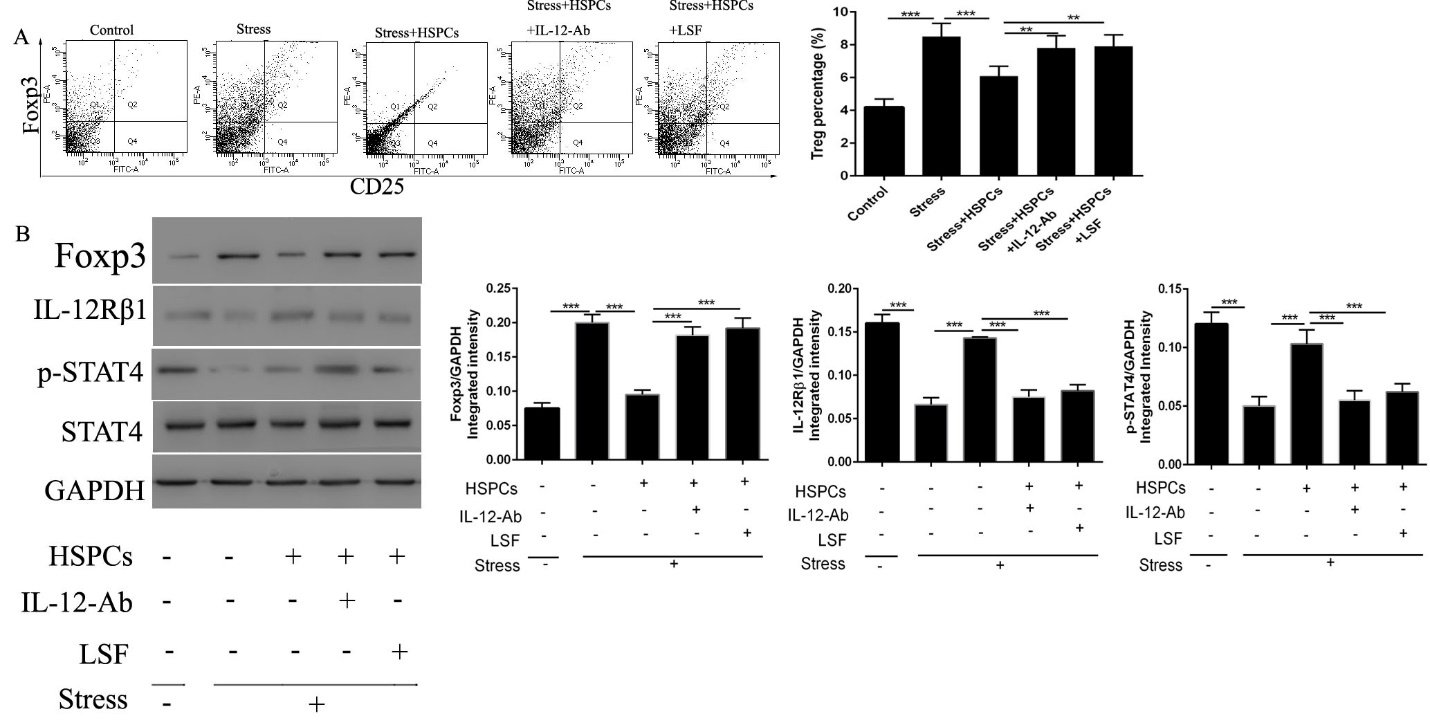
Figure 3: The HSPC-mediated protective effects against the proliferation of Tregs induced by chronic stress is dependent on IL-12/STAT4 signaling. BALB/c male mice (6-8 weeks old) were injected with CD34+HSPCs (1x106) in 100µl PBS with or without the IL-12 neutralization antibody or the selective STAT4 inhibitor LSF immediately before the initiation of restraint stress. (A) Inhibition of the IL-12/STAT4 signaling pathway abolished the protective effects of HSPCs. The isolated CD4+ cells in spleens were stained with PE-FOXP3 and FITC-CD25. (B) The protein expression levels of FOXP3, IL-12Rβ1, and p-STAT4 in the spleens were determined by western blotting. n=5 per group. ***P<0.001. HSPC, hematopoietic stem progenitor cell; LSF, Lisofylline.
As shown in Figure 3A, Treg proliferation was induced in stressed mice following HSPCs + IL-12-Ab or LSF administration. The expression of FOXP3 exhibited the same alteration in expression with the frequency of Tregs (Figure 3B). The protective effects of HPSCs were abolished following treatment with the neutralization of IL-12 or the inhibition of STAT4. We also examined whether HPSCs promoted the activation of IL-12/STAT4 signaling. Interestingly, HSPCs upregulated the expression of IL-12Rβ1 and p-STAT4 (Figure 3B), suggesting that the protective effect of HSPCs is dependent on IL-12/STAT4 signaling.
HSPCs Stimulate the Production of IL-12 by Splenocytes in Chronic Restraint Stress
IL-12 inhibited tumor-induced Treg proliferation in vitro [17, 19]. Injection of HSPCs inhibited the frequency of Tregs by promoting the production of IL-12. Thus, how HSPCs stimulated the production of IL-12 to play a protective effect on the proliferation of Tregs in chronic stress was next determined. Whether IL-12 was produced from the injected HSPCs or the splenocytes was determined. HSPCs were cultured in vitro and the IL-12 content in the supernatant was determined. HSPCs cannot secrete IL-12 under normal conditions (Figure 4A). Thus, we examined the role of mouse serum supplementation on IL-12 production by HSPCs. Neither serum from control mice nor stressed mice promoted the production of IL-12 by HSPCs (Figure 4B). As HSPCs did not produce IL-12 themselves, it was speculated that HSPCs may stimulate IL-12 production from splenic cells. To verify this, splenocytes were collected from stressed mice, cultured for 12 h and labeled for intracellular IL-12 and analyzed by FCM. The stressed mice treated with HSPCs had a significantly higher proportion of IL-12-expressing splenocytes than stressed mice without HSPC treatment (Figure 4C). These data revealed that HSPCs stimulated the production of IL-12 by splenic cells during chronic stress.
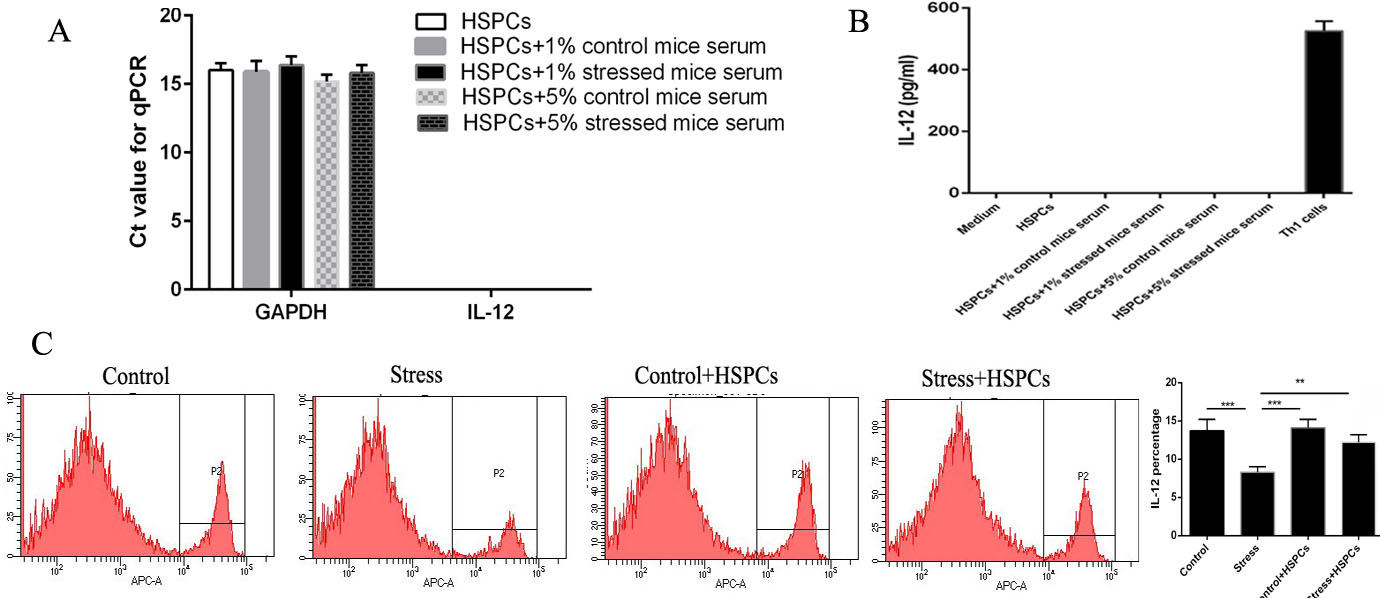
Figure 4: HSPCs stimulate the production of IL-12 in splenocytes. (A and B) CD34+HSPCs were cultured with serum from control or stressed mice. Supernatants and cells were harvested after 24 h. IL-12 expression in the supernatants were determined by ELISA. IL-12 mRNA expression was in HSPCs was examined. (C) The proportion of splenocytes from stressed mice treated with or without CD34+HSPCs were determined using flow cytometry. n=3 per group. ***P<0.001. HSPC, hematopoietic stem progenitor cell.
Discussion
Chronic Stress can disturb behavioral and endocrine homeostasis, alter the immunological response as a ubiquitous burden of modern-day life [20]. Our previous study revealed that HSPCs alleviated lymphocyte apoptosis induced by chronic stress [10]. In this study, HSPCs were shown to prevent Treg proliferation in a mouse model of chronic stress, and the molecular mechanism was related to the production of IL-12 after HSPC treatment.
IL-12 mobilizes HSPCs to the periphery and promotes hematopoiesis in vivo [21, 22]. IL-12 inhibits the expression of FOXP3 and the proliferation of Tregs by promoting IFN-γ, which mediated the inhibition of the proliferation of Tregs [17, 18]. Chronic stress impaired immune functions, as is indicated by the reduced levels of Th1 cytokines, including IFN-γ and IL-12 [23]. In vivo studies showed that certain stress-associated conditions such as social confrontation, wet-cage exposure, surgery, and the administration of corticosterone, epinephrine, or prostaglandin-E2 could inhibit the production of IL-12 [3, 24]. Our previous study revealed that chronic restraint stress induced the reduction of IL-12 [3]. The mechanisms of IL-12 suppression may be related to apoptosis of effector T cells [10], suppression of Th1 indices in the stress [25]. Chronic stress induced the production of glucocorticoids to inhibit Th1 immune responses, which selectively suppressed IL-12/STAT4 signaling in T lymphocytes, to inhibit the Th1 cellular immune response and cause a shift toward the Th2 humoral immune response [26].
IL-12 treatment inhibited the proliferation of Tregs by activating the IL-12 receptor, which reduced the production of IL-2, resulting in IFN-γ independent suppression of Tregs, as IL-2 is required for their survival and expansion [17]. IFN-γ signaling directly leads to cell cycle arrest in Tregs, and the IL-12/IFN-γ axis suppressed tumor-induced proliferation and expansion of Tregs in vitro [17]. Previous work showed that IL-12 reversed the suppressive function mediated by Tregs and IL-12 promoted the alteration of Tregs into IFN-γ-expressing cells [27]. These studies implied that chronic stress-induced Treg proliferation was related to a reduction in IL-12 expression. rIL-12 treatment before the initiation of stress blocked the proliferation of Tregs, and rIL-12 diminished the differentiation of Tregs in splenocytes stimulated with TGF-β1 in vitro, suggesting that IL-12 plays a critical role in the maintenance of the equilibrium of Tregs. IL-12 has been indicated to exert an effect on HSPCs by stimulating hematopoiesis recovery [21]. A low dose of IL-12 has been shown to play a protective role in hematopoiesis, which attenuates severe myelosuppression [22]. The potent immunological and multilineage hematopoietic effects triggered by IL-12 support the notion that rIL-12 may be used as a medical countermeasure and adjuvant immunotherapy for immune impairment induced by chronic stress.
Injection of HSPCs in chronic-stressed mice reversed the reduction in IL-12 levels and suppressed the proliferation of Tregs. However, neutralization of IL-12 or use of a selective STAT4 inhibitor abrogated the protective effects of HSPCs on Tregs, which suggested that HSPCs improved the impaired immune outcomes by promoting the production of IL-12 in chronic stress.
HSPCs promoted the production of IL-12 in stressed mice; injection of HSPCs alone (without stress) did not increase the production of IL-12 by splenocytes. Thus, the increase in IL-12 levels required both the presence of both stress and HSPCs. The underlying mechanisms by which high serum levels of IL-12 after HSPC treatment in chronic stress are achieved requires further study.
Conclusion
In conclusion, this study revealed that HSPCs diminished the proliferation and suppressive effects of Tregs induced by chronic restraint stress via IL-12/STAT4 signal and counteracted chronic stress induced immune dysfunction.
Funding
This work was supported by the hatching funding of Renmin Hospital of Wuhan University granted to Dr. Haiju Zhang (grant number: RMFH0001).
- Author Contributions: HZ designed the study and wrote the paper; HZ, SY and BY contributed to the experiments and the data analysis. All authors read and approved the paper.
- Data Availability: The datasets used and/or analyzed of the study are available from the corresponding author on reasonable request.
- Declarations Conflict of interest: The authors have no relevant interests to disclose.
- Ethical Approval: This study was approved by the Ethics Committee of Renmin Hospital of Wuhan University.
References
-
Dhabhar FS (2014) Effects of stress on immune function: the good, the bad, and the beautiful. Immunol Res 58(2- 3): 193-210.
-
Househam AM, Peterson CT, Mills PJ, Chopra D (2017) The effects of stress and meditation on the immune system, human microbiota, and epigenetics. Adv Mind Body Med 31(4): 10-25.
-
Hu D, Wan L, Chen M, Caudle Y, LeSage G, et al. (2014) Essential role of IL-10/STAT3 in chronic stress-induced immune suppression. Brain Behav Immun 36: 118-127.
-
Zhang H, Caudle Y, Wheeler C, Zhou Y, Stuart C, et al. (2018) TGF-β1/Smad2/3/Foxp3 signaling is required for chronic stress-induced immune suppression. J Neuroimmunol 314: 30-41.
-
Xiang Y, Yan H, Zhou J, Zhang Q, Hanley G, et al. (2015) The role of toll-like receptor 9 in chronic stress-induced apoptosis in macrophage. PLoS One10: e0123447.
-
Jin J, Wang X, Wang Q, Guo X, Cao J, et al. (2013) Chronic psychological stress induces the accumulation of myeloid-derived suppressor cells in mice. PLoS One 8(9): e74497.
-
Kim HR, Moon S, Lee HK, Kang JL, Oh S, et al. (2012) Immune dysregulation in chronic stress: a quantitative and functional assessment of regulatory T cells. Neuroimmunomodulation 19(3): 187-194.
-
Krause DS, Ito T, Fackler MJ, Smith OM, Collector MI, et al. (1994) Characterization of murine CD34, a marker for hematopoietic progenitor and stem cells. Blood 84(3): 691-701.
-
Laura B, Donald AF, Deling Y, Lesage GD, McCall CE, et al. (2012) Hematopoietic stem-progenitor cells restore immunoreactivity and improve survival in late sepsis. Infect Immun 80(2): 602-611.
-
Zhou Y, Li H, Siddiqui N, Caudle Y, Zhang H, et al. (2017) Hematopoietic stem progenitor cells prevent chronic stress-induced lymphocyte apoptosis. J Neuroimmunol 309: 72-76.
-
Heinrichs J, Bastian D, Veerapathran A, Anasetti C, Betts B, et al. (2016) Regulatory T-cell Therapy for graft- versus-host disease. J Immunol Res Ther 1(1): 1-14.
-
Cao G, Yang Q, Zhang S, Xu C, Roberts AI, et al. (2014) Mesenchymal stem cells prevent restraint stress- induced lymphocyte depletion via interleukin-4. Brain Behav Immun 38: 125-132.
-
Hajishengallis G, Shakhatreh MA, Wang M, Liang S (2007) Complement receptor 3 blockade promotes IL- 12-mediated clearance of Porphyromonas gingivalis and negates its virulence in vivo. J Immunol 179(4): 2359- 2367.
-
Liang S, Krauss JL, Domon H, McIntosh ML, Hosur KB, et al. (2011) The C5a receptor impairs IL-12-dependent clearance of Porphyromonas gingivalis and is required for induction of periodontal bone loss. J Immunol 186(2): 869-877.
-
Yang Z, Chen M, Fialkow LB, Ellett JD, Wu R, et al. (2003) Inhibition of STAT4 activation by lisofylline is associated with the protection of autoimmune diabetes. Ann N Y Acad Sci 1005: 409-411.
-
Lim WC, Olding M, Healy E, Millar TM (2018) Human endothelial cells modulate CD4+ T cell populations and enhance regulatory T cell suppressive capacity. Front Immunol 9: 565.
-
Cao X, Leonard K, Collins LI, Cai SF, Mayer JC, et al. (2009) Interleukin 12 stimulates IFN-gamma- mediated inhibition of tumor-induced regulatory T-cell proliferation and enhances tumor clearance. Cancer Res 69(22): 8700-8709.
-
Tarique M, Saini C, Naqvi RA, Khanna N, Sharma A, et al. (2017) IL-12 and IL-23 modulate plasticity of FoxP3+ regulatory T cells in human Leprosy. Mol Immunol 83: 72-81.
-
Zhao J, Zhao J, Perlman S (2012) Differential effects of IL- 12 on Tregs and non-Treg T cells: roles of IFN-γ, IL-2 and IL-2R. PLoS One 7(9): e46241.
-
Silberman DM, Wald MR, Genaro AM (2003) Acute and chronic stress exert opposing effects on antibody responses associated with changes in stress hormone regulation of T-lymphocyte reactivity. J Neuroimmunol 144(1-2): 53-60.
-
Jackson JD, Yan Y, Brunda MJ, Kelsey LS, Talmadge JE (1995) Interleukin-12 enhances peripheral hematopoiesis in vivo. Blood 85(9): 2371-2376.
-
Chen T, Burke KA, Zhan Y, Wang X, Shibata D, et al. (2007) IL-12 facilitates both the recovery of endogenous hematopoiesis and the engraftment of stem cells after ionizing radiation. Exp Hematol 35(2): 203-213.
-
Saul AN, Oberyszyn TM, Daugherty C, Kusewitt D, Jones S, et al. (2005) Chronic stress and susceptibility to skin cancer. J Natl Cancer Inst 97(23): 1760-1767.
-
Sorski L, Reznick M, Page GG, Ben Eliyahu S (2012) In vivo suppression of plasma IL-12 levels by acute and chronic stress paradigms: potential mediating mechanisms and sex differences. Brain Behav Immun 26(6): 996-1005.
-
Sarjan HN, Yajurvedi HN (2018) Chronic stress induced duration dependent alterations in immune system and their reversibility in rats. Immunol Lett 197: 31-43.
-
Franchimont D, Galon J, Gadina M, Visconti R, Zhou Y, et al. (2000) Inhibition of Th1 immune response by glucocorticoids: dexamethasone selectively inhibits IL- 12-induced Stat4 phosphorylation in T lymphocytes. J Immunol 164(4): 1768-1774.
-
Feng T, Cao AT, Weaver CT, Elson CO, Cong Y (2011) Interleukin-12 converts Foxp3+ regulatory T cells to interferon-γ-producing Foxp3+ T cells that inhibit colitis. Gastroenterology 140(7): 2031-2043.
- Origin, Evolution, and Functional Impact of Short Insertion- Deletion Variants in Human Genomes: A Review
- Harnessing Molecular Glues for Next-Generation Vaccine, Cancer and Cardiovascular Disease Drug Development: A Comprehensive Review
- Lateral Cervical Epidermal Inclusion Cyst in a Paediatric Patient: A Rare Case Report
- Malarial Plasmodium Falciparum with Hepatitis B and C Virus Infections among Blood Donors in Ife Central Local Government Area, Ile Ife, Osun State, Nigeria
- Withanolides and Withaferin A- What’s next in Ashwagandha Research
- Designing of Dual Pulse Photoacoustic Tomography for Imaging of Drug-Response and Tumor Growth