Unveiling of Potential Zika Virus RNA polymerase Inhibitors Using Structure based Virtual Screening
Zika virus (ZIKV) infection has been associated with Guillain-Barre syndrome in adults and Microcephaly in infants. However, there is no currently approved antiviral drug against ZIKV. The 103-kDa NS5 protein is the largest ZIKV protein whose C-terminal portion has RNA-dependent RNA polymerase activity and N-terminal RNA cap-processing activity (methyltransferase domain). Therefore, it is an important target for synthesis/discovery of novel anti-ZIKV drugs. A database consisting of 720 marine and plant derived compounds of South Africa was used for virtual screening against a potential inhibitor binding site of ZIKV’s NS5 protein. All its cataloged compounds were screened for their drug likeliness using Lipinski’s parameters. Thereafter, 200 selected compounds were used for virtual screening for their affinities for the target site. Finally, top 5 ligands were redocked to get a detailed insight into their interaction with conserved amino acids of the target site. An additional screening was mounted to filter out the potential toxic compounds among the top 5 likely candidates by AdmetSAR web-server. Two lead compounds, SANC00235 and SANC00257 fulfilled all the criteria laid down by us. These compounds can be potential candidates for further in vitro and in vivo studies in order to validate their anti-ZIKV activity.
Introduction
In 1947, Zika virus (ZIKV) was detected for the first time in a sentinel rhesus monkey in the Zika forest of Uganda [1]. After 21 years of its detection, the first human infected with ZIKV was in Nigeria [2]. For decades ZIKV remained relatively dormant, infecting mainly monkeys and occasionally causing a mild disease in humans residing across a narrow equatorial belt in Africa and Asia [3]. ZIKV infection has spread out of its epicenter after 2007. Pacific islands, including Federate States of Micronesia were among the first regions to face its outbreak. More Pacific islanders were infected in 2013-2014. In 2014, Americas were also included in the list of endemic countries [3, 4]. Till date, More than 60 countries including 23 in Latin American and the Caribbean countries have been affected with ZIKV [5, 6, 7]. Zika virus infection in humans can cause maculopapular rash, arthralgia, conjunctivitis, myalgia and headache [8]. Recently, it was found that ZIKV can also lead to some serious conditions like, Guillain-Barre syndrome in adults and microcephaly in new born infants [9, 10]. Therefore, in 2015, the World Health Organization (WHO) declared ZIKV as a public health emergency [11]. Moreover, ZIKV infection can induce testicular and epididymal damage, causing cell death and destruction of the seminiferous tubules [12]. It can become a long‐term persistent infection and can also be sexually transmitted [13, 14]. Unfortunately, not a single drug or vaccine is available for ZIKV treatment or prevention, which is of a grave concern ZIKV belongs to flavivirus family that also includes Yellow fever virus, West Nile virus, Spondweni virus, Japanese encephalitis virus and Dengue virus [15]. Zika virus has a 11Kb positive single strand RNA genome for encoding a polyprotein, which is cleaved proteolytically yielding three structural proteins, i.e. envelope (E), membrane precursor (PrM), and capsid (C) proteins, and seven non-structural proteins namely NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5 [16].
103-kDa NS5 is the largest, nonstructural and most conserved protein containing an N-terminal methyltransferase (MTase) domain for viral RNA capping and a RNA-dependent RNA polymerase (RdRp) domain at C-terminal responsible for viral RNA synthesis [14]. Both these domains together cooperate in initiation and elongation during viral RNA synthesis [11, 17]. Moreover, NS5 protein can also inhibit type I interferon (IFN) signaling to escape antiviral defense in the host [18]. The essential role of NS5 in viral replication and immunosuppression makes it an ideal target for antivirals [19]. A recent report has suggested a potential target site termed as ‘N-pocket’ near active site in RdRp domain of NS5 protein of ZIKV [19, 20]. This N-pocket is conserved in both zika and dengue virus and plays an important role in RNA initiation and elongation (Figure 1). The main residues in N-pocket are Leu-513, Glu- 514, Leu-516, Cys-711, His-713, Arg-731, Arg-739, Met-763, Trp-805, His-800 and Gly-801. RNA tunnel, through which RNA enters for replication is also located close to this pocket [21]. Therefore, targeting this site can effectively halt viral replication and proliferation [19].
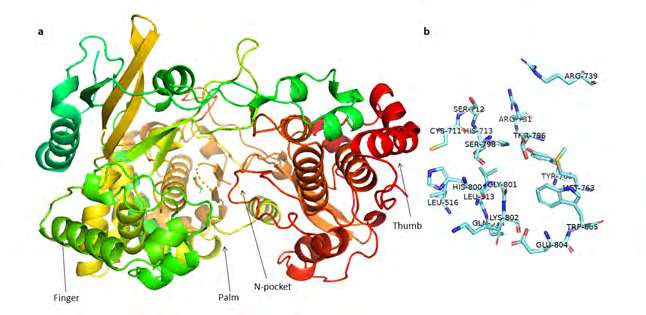
Natural products are among the most successful sources of potential drug candidates [22]. Their structures have high chemical diversity, biochemical specificity and other molecular properties which make them better candidates for drug discovery than synthetic and combinatorial compounds [23]. Even today, less than 10% of the total world biodiversity has been evaluated for their potential biological activity, hence many more compounds await their test for real therapeutic value [23]. Structure based virtual screening is one of the most widely used techniques to filter out potential candidates from a huge library/database. Therefore, we selected a database of natural compounds found in South Africa in order to screen them for their ability to inhibit the N-pocket of NS5 protein because South Africa is one of the few countries where zika virus infection is scarce [24, 25].
Material and Methods
Ligands and Protein Preparation
A database of South African natural compounds was employed for this study [24]. Energy minimized structures were downloaded for further studies. X-ray structure file of NS5 protein with ID 5TFR was obtained from protein databank and Het atoms were deleted [26]. Structure of NS5 was optimized using steepest descent algorithm of Swiss-pdb Viewer software [27]. Polar hydrogen atoms and gasteiger charges were added to the protein.
Screening for Drug-Likeness
All ligands obtained from the database were screened for their drug-likeness potential. DruLi To software was used to filter compounds on the basis of Lipinski’s rule of five [28, 29]. Compounds selected as potential drug molecules were shortlisted for further studies.
Structure based Virtual Screening
Structure based virtual screening is one of the most effective and efficient methods for filtering compounds from huge library. PyRx software was used for structure based virtual screening that employed Autodock Vina and Autodock tools for molecular docking [30, 31, 32]. Vina
module was selected for virtual screening as it is fast and recommended for screening large structure libraries. Grids was adjusted engulfing the target residues in the N-pocket of NS5 protein with its center at x=18.812 y=-53.14 z=1.346. Top 5 compounds were short listed based on their binding affinity for target site.
Re-docking and ADMET studies
Lead compounds were re-docked using Autodock tools to get a detailed insight into the interaction of compounds with residues of target site. Autodock is one of the most widely used software for molecular docking. It predicts and calculates the interaction between ligands and macromolecules along with their binding energies, number of hydrogen bonds and Van der Waal energies. A grid box was generated around N-pocket with center at 21 x -55 x 0.24. In order to magnify the cluster difference, 100 GA runs were performed for each selected compound. Pymol and Schrodinger Mestro softwares were used to analyze the docked poses [33, 34]. Moreover, ADMETsar web server was used to predict ADMET (Absorption, Distribution, Metabolism, Excretion and Toxicity) properties of selected compounds [35]. ADMET property predictions play a crucial role in drug discovery because majority of drugs get rejected on the basis of these parameters.
Results and Discussion
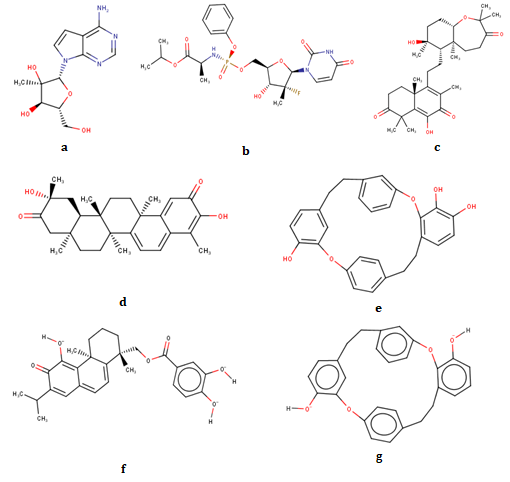
Virtual screening (VS) provides an efficient platform to filter out ligands of potential/interest from a huge library or database. All natural compounds from South African database consisting of 720 compounds were downloaded to test their potential to inhibit NS5 protein of ZIKV [14, 24]. Before proceeding for VS, compounds were screened for their drug likeness potential. Lipinski’s rule of five was employed as it is an important and most widely used parameter in drug discovery [28]. 200 out of 720 compounds passed Lipinski’s rule. Thereafter, virtual screening of these 200 shortlisted compounds was done in order to predict their binding affinities for pocket of interest i.e. N-pocket of NS5 protein. The binding energies of these selected compounds obtained in VS are shown in Table 1. Top 5 compounds on the basis of their binding affinity towards target site were selected. Figure 2 shows chemical structures of selected and template compounds.
| S No | Compound/ Database ID | Binding affinity (kcal/mol) | No. of clusters | H-Bonds | Hydrophobic interaction | ||
|---|---|---|---|---|---|---|---|
| Vina | Auto-dock | No. | Interacting residues | ||||
| 1 | 7DMA | -7.2 | -5.01 | 40 | 3 | Asp-665, Asp-666, Ile-799 | Tyr-609 |
| 2 | Sofosbuvir | -7.91 | -4.63 | 3 | 2 | Tyr-609, Asn-612 | Ile-799 |
| 3 | SANC00772 | -9.95 | -7.4 | 85 | 1 | Lys-462 | Trp-797 |
| 4 | SANC00235 | -9.78 | -9.08 | 33 | 3 | Trp-797, Tyr-609, Asn-612 | Tyr-609, Ile-799 |
| 5 | SANC00257 | -9.75 | -8.37 | 100 | 2 | Tyr-609, Val-607 | Tyr-609, Ile-799 |
| 6 | SANC00373 | -9.74 | -9.72 | 29 | 5 | Lys-462, Asp-373, Ile-799 | Lys-461, Arg-739, Tyr- 609, Thr-796, Trp-797, Ile-799 |
| 7 | SANC00768 | -9.71 | -7.35 | 77 | 1 | Lys-462 | Trp-797 |
Table 1: Details of binding energy, number of clusters and H-bonds of template and selected compounds.
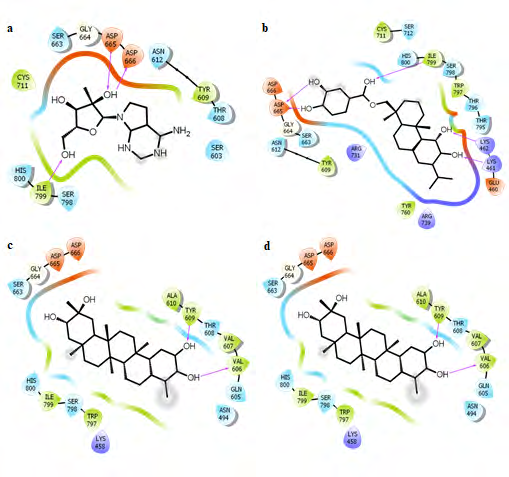
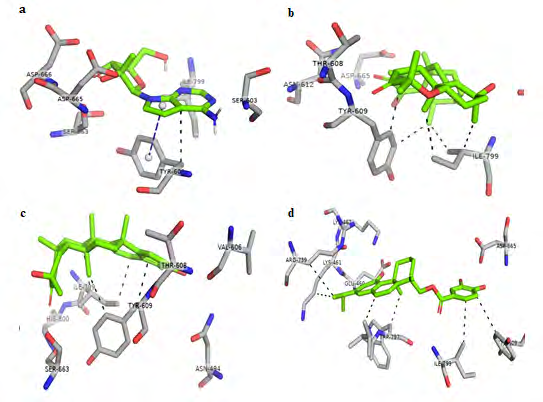
To get a detailed insight of interaction between selected compounds and residues of N-pocket, shortlisted compounds were re-docked using autodock software [31]. The compound cataloged as SANC00373 was found to have maximum binding affinity i.e. -9.72 Kcal mol-1. It forms H-bonds with Lys-461, Lys-462, Ile-799 and Asp-665 (Figure 3). Moreover, hydrophobic interactions with Lys-461, Tyr-609, Arg- 739, Thr-796, Trp-797 and Ile-799 were also involved in the binding of the compound with N-pocket (Figure 4). The compound cataloged as SANC00235 ranked second when re-docked with a binding energy of -9.08 Kcal mol-1.
It forms H-bonds with Asp-665 and Ile-799. Moreover, Tyr- 609 and Ile-799 residues develop hydrophobic interactions with this compound resulting in a strong binding. Similarly, SANC00257 binds with an affinity of -8.37 kcal mol-1, forming H-bonds with Val-606 and Tyr-609 along with hydrophobic interactions with Tyr-609 and Ile-799. Moreover, the compounds SANC00772 and SANC00768 bind quite strongly with the binding energies of -7.4 Kcal mol-1 and -7.35 Kcal mol-1, respectively. Both of them form single H-bond with Lys-462 and hydrophobic interaction with Trp-797.
For prediction purpose, ADMET profiles were obtained using ADMETsar web server as shown in Table 2. Two compounds i.e. SANC00235 and SANC00257, were found to be safe on the basis of various ADMET parameters. They are capable of crossing the blood brain barrier and their intestinal absorption also seems to be good [36].
| ADMET parameters | SANC00772 | SANC00235 | SANC00257 | SANC00373 | SANC00768 |
|---|---|---|---|---|---|
| Blood-Brain Barrier | BBB+ | BBB+ | BBB+ | BBB- | BBB+ |
| Human Intestinal Absorption | HIA+ | HIA+ | HIA+ | HIA+ | HIA+ |
| Caco-2 Permeability | impermeable | permeable | permeable | permeable | permeable |
| Renal Organic Cation Transporter | non-inhibitor | non-inhibitor | non-inhibitor | non-inhibitor | non-inhibitor |
| CYP450 1A2 Inhibitor | inhibitor | non-inhibitor | non-inhibitor | inhibitor | inhibitor |
| CYP450 2C9 Inhibitor | inhibitor | non-inhibitor | non-inhibitor | non-inhibitor | inhibitor |
| CYP450 2D6 Inhibitor | non-inhibitor | non-inhibitor | non-inhibitor | non-inhibitor | non-inhibitor |
| CYP450 2C19 Inhibitor | non-inhibitor | non-inhibitor | non-inhibitor | non-inhibitor | inhibitor |
| CYP450 3A4 Inhibitor | non-inhibitor | non-inhibitor | non-inhibitor | non-inhibitor | non-inhibitor |
| AMES Toxicity | non-Ames toxic | non-Ames toxic | non-Ames toxic | non-Ames toxic | non-Ames toxic |
| Carcinogens | non- carcinogens | non- carcinogens | non-carcinogens | non-carcinogens | non-carcinogens |
| Acute Oral Toxicity (mg/kg) | >500 | >500 | >5000 | >500 | >500 |
| <5000 | <5000 | <5000 | <5000 | ||
| Aqueous solubility (LogS) | -2.1523 | -4.1537 | -4.0769 | -5.029 | -2.3566 |
Table 2: ADMET parameters of the top 5 selected lead compounds. ADMET results were generated using ADMETsar server. Red color den
Moreover, they were also found to be non-mutagenic and non-carcinogenic. Similarly, their acute oral toxicity was predicted to be only at relatively higher doses. Besides the above, their aqueous solubility were favorable for a drug molecule. SANC00772 was predicted to be impermeable in Caco-2 cell lines of our in silico study, a parameter widely studied as a drug absorption test [37]. It was also assumed to be a CYP450 1A2 and CYP450 2C9 inhibitor. The compound cataloged as SANC00373 was unable to cross blood brain barrier, its water solubility was not promising too. Similarly, SANC00768 was predicted to act as an inhibitor for CYP450 1A2, CYP450 2C9 and CYP450 2C19. Therefore, the remaining two compounds i.e. SANC00235 and SANC00257, were safe based on the established criteria and can serve as potential candidates for anti-zika virus therapy.
Conclusion
NS5 is the most conserved protein of Zika virus and plays an important role in virus replication. Especially, N-pocket of NS5 protein is a promising domain to inhibit virus replication according to available literature. Therefore, it was selected as binding target for this in silico study. A total of 720 natural compounds from South Africa were screened for their appreciable binding affinity towards the said target site. Another filter was mounted on 200 compounds cleared as above for their ADMET characteristics. In a nutshell, two compounds cataloged as SANC00235 and SANC00257 exhibited outstanding binding affinities with maximum binding energies and significant number of clusters. Both the compounds followed Lipinski’s rule for drug likeness. Furthermore, they show promise when tested on ADMET parameters, suggesting them to be potential candidates for anti-ZIKV drug. However, further in vitro and in vivo studies are required to establish their actual therapeutic potential.
Conflict of Interest
Authors declare that there is no conflict of interest of any kind.
Acknowledgement
No funding was provided by any agency or institute for this work. Authors acknowledge department of biochemistry, Aligarh Muslim University.
References
-
Dick GWA (1952) Zika Virus (I). Isolations and serological specificity. Trans R Soc Trop Med Hyg 46(5): 509-520.
-
Moore DL, Causey OR, Carey DE (1975) Arthropod-borne viral infections of man in nigeria, 1964-1970. Ann Trop Med Parasitol 69(1): 49-64.
-
Fauci AS, Morens DM (2016) Zika Virus in the America- Yet Another Arbovirus Threat. N Engl J Med 374(7): 601- 604.
-
Musso D, Nilles EJ, Cao-Lormeau VM (2014) Rapid spread of emerging Zika virus in the Pacific area. Clin Microbiol Infect 20(10): O595-O596.
-
Gatherer D, Kohl A (2016) Zika virus: A previously slow pandemic spreads rapidly through the Americas. J Gen Virol 97(2): 269-273.
-
Duffy MR, Chen TH, Hancock WT, Powers AM, Kool JL, et al. (2009) Zika virus outbreak on Yap Island, Federated States of Micronesia. N Engl J Med 360(24): 2536-2543.
-
Faria NR, Do Socorro Da Silva Azevedo R, Kraemer MUG, Souza R, Cunha MS, et al. (2016) Zika virus in the Americas: Early epidemiological and genetic findings. Science 352(6283): 345-349.
-
Simpson DIH (1964) Zika virus infection in man. Trans R Soc Trop Med Hyg 58(4): 335-338.
-
Rasmussen SA, Jamieson DJ, Honein MA, Petersen LR (2016) Zika Virus and Birth Defects-Reviewing the Evidence for Causality. N Engl J Med 374: 1981-1987.
-
Cao-Lormeau VM, Blake A, Mons S, Lastere S, Roche C, et al. (2016) Guillain-Barre Syndrome outbreak associated with Zika virus infection in French Polynesia: A case- control study. Lancet 387(10027): 1531-1539.
-
Saiz JC, Vázquez-Calvo Á, Blázquez AB, Merino-Ramos T, Escribano-Romero E, et al. (2016) Zika Virus: the Latest Newcomer. Front Microbiol 7: 496.
-
Govero J, Esakky P, Scheaffer SM, Fernandez E, Drury A, et al. (2016) Zika virus infection damages the testes in mice. Nature 540: 438-442.
-
Mansuy JM, Dutertre M, Mengelle C (2016) Zika virus: High infectious viral load in semen, a new sexually transmitted pathogen? Lancet Infect Dis 16(4): 405.
-
Duan W, Song H, Wang H, Chai Y, Su C, et al. (2017) The crystal structure of Zika virus NS5 reveals conserved drug targets. EMBO J 36(7): 919-933.
-
Vasudevan J, Skandhan A, Skandhan AKP (2018) Zika virus. Rev Med Microbiol 29(3): 43-50.
-
Faye O, Freire CCM, Iamarino A (2014) Molecular evolution of Zika Virus during its emergence in the 20th Century. PLoS Negl Trop Dis 8: 36.
-
Potisopon S, Priet S, Collet A, Decroly E, Canard B, et al. (2014) The methyltransferase domain of dengue virus protein NS5 ensures efficient RNA synthesis initiation and elongation by the polymerase domain. Nucleic Acids Res 42(18): 11642-11656.
-
Best SM, Morris KL, Shannon JG, Robertson SJ, Mitzel DN, et al. (2005) Inhibition of Interferon-Stimulated JAK-STAT Signaling by a Tick-Borne Flavivirus and Identification of NS5 as an Interferon Antagonist. J Virol 79(20): 12828-12839.
-
Wang B, Tan XF, Thurmond S, Zhang ZM, Lin A, et al. (2017) The structure of Zika virus NS5 reveals a conserved domain conformation. Nat Commun 8: 14763.
-
Lim SP, Noble CG, Seh CC, Soh ST, El Sahili A, et al. (2016) Potent Allosteric Dengue Virus NS5 Polymerase Inhibitors: Mechanism of Action and Resistance Profiling. PLOS Pathog 12(8).
-
Niyomrattanakit P, Chen YL, Dong H, Yin Z, Qing M, et al. (2010) Inhibition of Dengue Virus Polymerase by Blocking of the RNA Tunnel. J Virol 84(11): 5678-5686.
-
Dias DA, Urban S, Roessner U (2012) A Historical overview of natural products in drug discovery. Metabolites 2(2): 303-336.
-
Karthikeyan M, Balasubramanian T, Khaleel MI, Sahl M, Rashifa P (2015) A Systematic Review on Medication Errors. Int J Drug Dev Res 7(4): 9-11.
-
Hatherley R, Brown DK, Musyoka TM, Penkler DL, Faya N, et al. (2015) SANCDB: A South African natural compound database. J. Cheminform 7: 29.
-
Braack L, Venter M (2016) Key information on Zika virus and South Africa.
-
Upadhyay AK, Cyr M, Longenecker K (2017) Crystal structure of full-length Zika virus NS5 protein reveals a conformation similar to Japanese encephalitis virus NS5. Acta Crystallogr Sect Struct Biol Commun 73(Pt 3): 116- 122.
-
Guex N, Peitsch MC (1997) SWISS-MODEL and the Swiss- Pdb Viewer: An environment for comparative protein modeling. Electrophoresis 18(15): 2714-2723.
-
Lipinski CA (2004) Lead- and drug-like compounds: The rule-of-five revolution. Drug Discov Today Technol 1(4): 337-341.
-
Tool DL (2010) Drug Likeness Tool (DruLiTo), pp: 1-9.
-
Trott O, Olson AJ (2010) Software news and update AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J Comput Chem 31(12): 455-461.
-
Morris GM, Huey R, Lindstrom W (2009) AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J Comput Chem 30(16): 2785-2791.
-
Dallakyan S, Olson AJ (2015) Small-molecule library screening by docking with PyRx. Methods Mol Biol 1263: 243–250.
-
(2018) The PyMOL Molecular Graphics System, Version 2.0 Schrödinger, LLC.
-
Maestro S (2018) Schrödinger. Schrödinger Release.
-
Cheng F, Li W, Zhou Y (2012) AdmetSAR: A comprehensive source and free tool for assessment of chemical ADMET properties. J Chem Inf Model 52(11): 3099-3105.
-
Hussain A, Verma CK (2017) Molecular docking and in silico admet study reveals 3-(5-{[4-(aminomethyl) piperidin-1-yl] methyl}-1h-indol-2-yl)-1h-indazole- 6-carbonitrile as a potential inhibitor of cancer osaka thyroid kinase. Biomed Res 28(13): 5805–5815.
-
Kumar KKV, Karnati S, Reddy MB (2010) Caco-2 cell lines in drug discovery- an updated perspective. J basic Clin Pharm 1(2): 63-69.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells