Perspectives of Toxicity Associated with Nanocarrier Systems
The utility and diversified applications of various nanocarrier systems have led to the development of a wide variety of formulations with smart properties. Although these formulations offer several advantages over traditional delivery systems such as site-specific, time-dependent and controlled delivery of the medicaments but unfortunately the toxicological behavior of these has remained unexplored. There are several reports in the literature that have described the significant toxicity in major organs of animals. This toxicity has majorly associated with the formation of reactive oxygen species (ROS), elevation/ reduction in biomarker levels, induction of apoptosis and several other molecular changes. In this short compilation, we have summarized some toxicity reports which have been based on pre-clinical evidences and attributed to multiple organs of animals. These include the kidney, heart, lungs, liver and GIT prominently. Also, we have made an attempt to highlight the mechanism of the reported toxicity along with the toxic dose. This compilation may be helpful to drug developers and researchers to understand these issues and to design newer strategies during formulation to bypass these complications.
Introduction
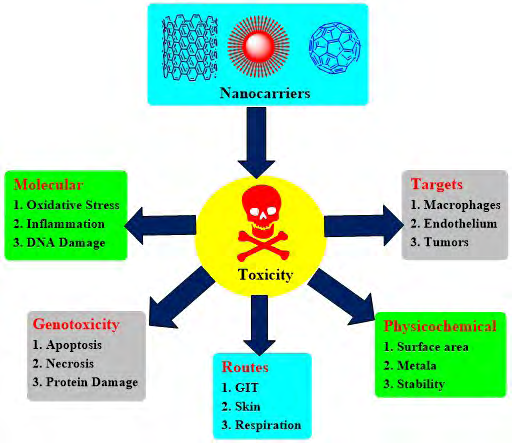
In the past decade, nanocarrier systems have been extensively explored for their divine drug delivery potentials and had been widely utilized in the development of targeted drug delivery systems, research and technology. Nanocarrier based drug delivery systems have been proven as blockbusters for site-specific delivery in the therapy of life-threatening ailments. In other words, we can coin these systems as smart nanocarriers because of their smart functionality and application in the development of smart drug delivery systems (SDDs). These delivery systems have bypassed the disadvantages of non-specific distribution and uncontrollable drug delivery patterns of traditional delivery systems. Smart nanocarrier systems include micelles, liposomes, dendrimers, carbon nanotubes (CNTs), multi-walled carbon nanotubes (MWCNTs), single-walled carbon nanotubes (SWCNTs) nanorods, nanoemulsions, phytosomes, magnetic nanoparticles, nanospheres, quantum dots and mesoporous silica nanoparticles (MSNs). Many nanocarrier-based formulations are already available in the market and some are in clinical trial phases. Despite several extraordinary advantages of these smart nanocarriers, there is a continuously emerging issue of toxicity of these systems. As a matter of great concern, continuous research is in progress specially focussing on toxicity and biocompatibility of nanocarriers. In general, nanoparticles are able to induce toxicity based upon their internalization site and composition. It is also revealed that nanoparticles can cause inflammation, oxidative stress and DNA damage [1]. Table 1 highlights some significant reports describing the potent toxicities exhibited by nanocarrier systems on different organs/parts of the body. Major consequences involved in nanocarrier toxicity in terms of targets, molecular involvement, genotoxicity, routes and physicochemical factors have been outlined in Figure 1.
| Carrier | Organ | Effects | Animal used /cell line | Mechanism of toxicity | Toxic Dose | Refer ence |
|---|---|---|---|---|---|---|
| CNTs | Lungs | Alteration in mitochondrial membrane potential | Rat alveolar macrophage cell line (NR8383) | Metal catalysed induction of reactive oxygen species (ROS) | 50 µg/mL | [2] |
| MWCNTs | Lungs | DNA damage | Female mice | Pulmonary inflammation induced by neutrophil influx in broncho-alveolar lavage (BAL)) and genotoxicity leading to DNA damage | 6-54 µg/ mL | [3] |
| SWCNTs | Lungs | Death due to blockage of airway | Male rats | Alveolar macrophage accumulation and lung tissue thickening | 5mg/Kg | [4] |
| MWCNTs | Lungs | Pulmonary lesion and collagen rich granuloma in the mice exposed | Guinea pigs (males) | Perivascular, peribronchial and interstitial permeation of inflammatory cells associated with central and peripheral atelectasis, emphysema and alveolar exudation | 1-5mg/Kg | [5] |
| SWCNTs | Heart | Progression of atherosclerosis | Mice | Aortic DNA damage | 10-40µg | [6] |
| CNTs | Foetus | The fetal and developmental abnormalities | Male and female mice | Increased resorptions during organogenesis, induction of oxidative stress due to ROS | 10mg/Kg | [7] |
| IONPs (Iron oxide nanoparticles) | Liver | Liver inflammation and necrosis | Adult male Wistar rats | Enhancement of free radicals and reduction of GSH in lung tissues | >2.2mg/Kg | [8] |
| Curcumin capped IONPs | Liver and kidney | Abnormal liver and kidney performance | Mice | Changes in the levels of biomarkers of liver and kidney | >5mg/Kg | [9] |
| Dendrimer coated IONPs | Liver | Edema and losing cytoplasm in the liver cells | Mice | Increase in blood urea nitrogen, bilirubin and histopathological abnormalities | 10mg/Kg | [10] |
| Platinum nanoparticles | Heart | Decrease in the heart rate, prolonged P-R intervals and finally complete A-V conduction block | Mice | Decrease in current densities of ion channels, conduction block and increased lactate dehydrogenase leak | 3-10mg/kg | [11] |
| CuO NPs (Copper oxide nanoparticles) | Liver and spleen. | Liver and kidney dysfunction | Female mice | Increased production of ROS leading to lymphocyte apoptosis | 100- 1000µg/Kg | [12] |
| CuO NPs | G.I.T. | G.I.T. Toxicity, an imbalance in antioxidant levels. | Artemia salina | Generation of oxidative stress and disturbances in antioxidant defence pathway | 12.2mg/L | [13] |
| TiO NPs 2 | Heart and liver | Heart injury and liver injury | Rat | Elevated reduced glutathione (GSH)/oxidized glutathione ratios due to increased plasma levels of glucose and GSH | 50-200mg/ Kg | [14] |
| TiO NPs 2 | Liver | Liver injury markers and a reduction in certain hematological parameters. | Female mice | Elevated levels of alanine aminotransferase, alkaline phosphatase, aspartate aminotransferase, lactate dehydrogenase and cholinesterase, total protein and the reduction total bilirubin, triglycerides, and the total cholesterol levels | 125- 250mg/Kg | [15] |
| TiO NPs 2 | Foetus | Fetal toxicity in pregnant mice | Mice | Elevated dopamine levels in the prefrontal cortex and neostriatum, abnormal fetal liver development | 0.25–1.00 mg/mL | [16] |
| Mesoporous Silica NPs | Kidney | Hemorrhage, vascular congestion, and renal tubular necrosis | Male mice | Renal tubular necrosis, vascular congestion in renal interstitium | 40mg/Kg | [17] |
| ZnO NPs | Fetus | Toxicity during gestation period | Rats | Multifocal mixed cell permeation, thrombosis in lung, tubular dilation in kidneys | 10-20mg/ Kg | [18] |
| Liposomes | Liver, Lung, Breast | Cytotoxicity | L 1210, HepG2, A549 cell lines | DNA damage due to the cationic surface charge | 0.25 µM P/ ml | [19] |
| Micells | Lung, Liver, Kidney | Polymeric micelle-based drug carriers trigger transient immunogenicity | Female Mice | Increased ROS production, Increase in cell volume | Dose dependent | [20] |
| Dendrimers | Lung, Liver, Kidney | Dendrimers, such as PPI, PAMAM, and PLL, exert significant in vitro cytotoxicity due to their surface catatonic groups | Mammalian Cells | High charge and strong interaction with the negatively charged cell membranes leading to destabilization and leakage and lysis of cytoplasmic proteins | Dose dependent | [21] |
Table 1: Reported toxicity of nanocarrier systems on various organs.
Possible Mechanisms of Toxicity
The various pre-clinical studies of nanocarrier systems have been carried out by several research groups including the toxicity along with its underlying mechanisms. A few significant mechanisms of toxicity revealed by various nanoparticles (NPs) have been described in the following sections.
Generation of Reactive Oxygen Species (ROS)
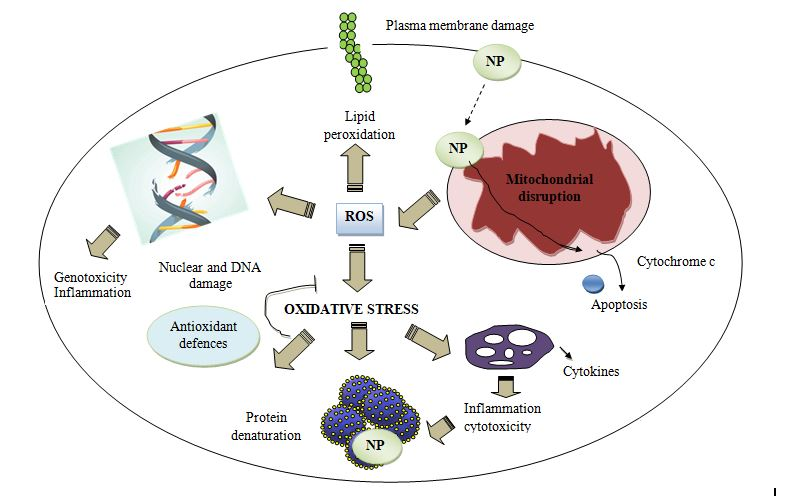
The physiological activity of nanocarriers leads to the generation of reactive oxygen species which include hydroxyl radicals, superoxide radicals as a result of activation of oxidative enzymes which ultimately is the prominent cause of oxidative stress (Figure 2) [22, 23, 24]. It is worth notable that the extent of this kind of stress has been reported majorly in nanocarriers systems possessing metals or impurities of transition metals [25, 26]. Deposition of nanoparticles in multiple organs leads to ROS generation and initiation of inflammation. This mechanism is not fully understood but it has been evidenced that oxidative stress affects intracellular calcium contents, transcription variables and induction of cytokines [27]. Elevated ROS adversely affect mitochondrial respiratory mechanisms and induces changes in protein structures in the endoplasmic reticulum and induce stress. These events lead to more production of ROS, severe DNA damage, induction of signals, more inflammatory events, cell death due to apoptosis and necrosis [28, 29].
The redox process may occur in the solution as well as on the nanoparticle surface leading to changes in the crystalline structure. Some nanocarrier preparations such as fullerenes, carbon dots, SWNTs and quantum dots produce ROS upon exposure to ultraviolet radiations or transition metals [30]. Exposure of a mother to titanium dioxide can cause changes in apoptotic genes and oxidative stress in the newborn offspring [31]. Nanoparticles possess a large surface area which can produce prominent ROS and leads to cytotoxicity. The CNS is highly sensitive towards oxidative stress due to abundant lipids, proteins, high oxygen consumption and weak antioxidant properties [32]. Therefore ROS causes maximum damage in CNS leading to neurodegenerative disorders and diseases. It has also been reported that nanoparticles are also capable of damaging dopaminergic neurons as a result of high production of ROS due to microglial stimulation [33].
Cellular uptake Mechanisms
The structural organization, chemistry and size of nanoparticles greatly influence the cellular entry, uptake and distribution of these systems. The cell membranes give entry to nanoparticles by the endocytosis process which is influenced by the nature, size and shape of the nanoparticles [34]. The size directly affects various cellular processes like target identification, circulatory residence time, concentration, uptake pathway and clearance. Smaller particles enter and exit with great ease; spherical particles get internalized inside the cells and negatively charged particles exhibit a low rate of endocytosis as compared to positively charged ones. Pinocytosis is the type of endocytosis that is meant for the intake of fluid or smaller solute particles whereas phagocytosis intakes the heavy and solid materials. Phagocytosis takes place through macrophages, neutrophils, monocytes and dendritic cells. Opposing to this, pinocytosis involves van der Waals, electrostatic, steric interactions and the formation of vesicles leading to free movement of nanoparticles between cells and multiple organelles [35, 36].
Followed by pinocytosis, the nanoparticles got located in various compartments of cells such as cell membrane, cytoplasm, mitochondria, lipid vesicles, nuclear membrane, nucleus and exert significant toxicity by causing organelle/ DNA damage leading to cell death [37, 38, 39]. Complications produced by nanoparticles upon localizing in particular organelles have been depicted in Figure 3. The shape of nanoparticles also affects the cellular uptake and has been reported highest in the case of nanorods in human cervical cancer cells followed by nanospheres, cylindrical and cubical shapes [40, 41]. Lysosomes are also a significant target for nanoparticle localization and toxicity due to endocytosis. NPs exert their toxicity in lysosomes due to cytoskeleton destruction, alkalization or overload [42].
![Figure 3: The shape of nanoparticles also affects the cellular uptake and has been reported highest in the case of nanorods in human cervical cancer cells followed by nanospheres, cylindrical and cubical shapes [40,41]. Lysosomes are also a significant target for nanoparticle localization and toxicity due to endocytosis. NPs exert their toxicity in lysosomes due to cytoskeleton destruction, alkalization or overload [42].](/fulltextimages/7731/fig_3.png)
Genotoxicity and Inflammation
The activation of microglia by nanoparticles leads to the initiation of inflammatory responses by secreting pro- inflammatory factors and ultimately causes cell dysfunction, death and cytotoxicity [43]. Nps have been also regarded as autophagy inducers with the potentials of inducing ROS- dependent and lysosome-dependent autophagy. Titanium, silicon, polymeric, oleic-acid coated nanoparticles are responsible for brain autophagy whereas zinc oxide NPs cause oxidative stress in macrophages leading to autophagy and apoptosis [44]. Nanoparticles of varying sizes accumulate in mitochondria and lead to abnormal electron transport chain mechanisms [45]. This oxidative stress ultimately leads to genotoxicity due to DNA modifications and cell injuries [46]. Epigenetic effects are also prominent in chromatin due to acetylation/methylation of histones, mutagenic DNA damage and abolition of DNA repair pathways which is the prominent cause of Ni-nanoparticles induced carcinomas [47, 48, 49, 50].
Conclusion
It is evident from the above reports that along with therapeutic efficacy the nanocarrier systems exhibit a significant amount of toxicity. This toxicity has been attributed to several factors like ROS generation, inflammation, endocytosis, nanoparticle size, shape and localization, etc. This is a matter of immense concern and researchers/drug developers should work in this direction so as to reduce the induced toxicity. Although there are a few approaches that have been successfully utilized for reducing the toxicity such as modification in size, shape, shell, surface charge and route of administration; still a keen work towards this direction is the demand of the hour.
References
-
Rizvi SA, Saleh AM (2018) Applications of nanoparticle systems in drug delivery technology. Saudi Pharmaceutical J 26(1): 64-70.
-
Pulskamp K, Diabate S, Krug HF (2007) Carbon nanotubes show no sign of acute toxicity but induce intracellular reactive oxygen species in dependence on contaminants. Toxicol lett 168(1): 58-74.
-
Poulsen SS, Jackson P, Kling K, Knudsen KB, Skaug V, et al. (2016) Multi-walled carbon nanotube physicochemical properties predict pulmonary inflammation and genotoxicity. Nanotoxicology 10(9): 1263-1275.
-
Warheit DB, Laurence BR, Reed KL, Roach DH, Reynolds GA, et al. (2004) Comparative pulmonary toxicity assessment of single-wall carbon nanotubes in rats. Toxicol sci 77(1): 117-125.
-
Grubek-Jaworska H, Nejman P, Czumińska K, Przybyłowski T, Huczko A, et al. (2006) Preliminary results on the pathogenic effects of intratracheal exposure to one-dimensional nanocarbons. Carbon 44(6): 1057-1063.
-
Li Z, Hulderman T, Salmen R, Chapman R, Leonard SS, et al. (2007) Cardiovascular effects of pulmonary exposure to single-wall carbon nanotubes. Environmental health perspectives 115(3): 377-82.
-
Philbrook NA, Walker VK, Afrooz AN, Saleh NB, Winn LM (2011) Investigating the effects of functionalized carbon nanotubes on reproduction and development in Drosophila melanogaster and CD-1 mice. Reproductive Toxicology 32(4): 442-448.
-
Almeida JP, Chen AL, Foster A, Drezek R (2011) In vivo biodistribution of nanoparticles. Nanomedicine 6(5): 815-835.
-
Elbialy NS, Aboushoushah SF, Alshammari WW (2019) Long-term biodistribution and toxicity of curcumin capped iron oxide nanoparticles after single-dose administration in mice. Life Sci 230: 76-83.
-
Salimi M, Sarkar S, Fathi S, Alizadeh AM, Saber R, et al. (2018) Biodistribution, pharmacokinetics, and toxicity of dendrimer-coated iron oxide nanoparticles in BALB/c mice. Int J Nanomedicine 13: 1483.
-
Lin CX, Gu JL, Cao JM (2019) The acute toxic effects of platinum nanoparticles on ion channels, transmembrane potentials of cardiomyocytes in vitro and heart rhythm in vivo in mice. Int J nanomedicine 14: 5595.
-
Dey A, Manna S, Adhikary J, Chattopadhyay S, De S, et al. (2019) Biodistribution and toxickinetic variances of chemical and green Copper oxide nanoparticles in vitro and in vivo. J Trace Elem Med Biol 55: 154-169.
-
Madhav MR, David SE, Kumar RS, Swathy JS, Bhuvaneshwari M, et al. (2017) Toxicity and accumulation of Copper oxide (CuO) nanoparticles in different life stages of Artemia salina. Environ Toxicol Pharmacol 52: 227-238.
-
Wang Y, Chen Z, Ba T, Pu J, Chen T, et al. (2013) Susceptibility of young and adult rats to the oral toxicity of titanium dioxide nanoparticles. Small 9(9‐10): 1742- 1752.
-
Duan Y, Liu J, Ma L, Li N, Liu H, et al. (2010) Toxicological characteristics of nanoparticulate anatase titanium dioxide in mice. Biomaterials 31(5): 894-899.
-
Naserzadeh P, Ghanbary F, Ashtari P, Seydi E, Ashtari K, et al. (2018) Biocompatibility assessment of titanium dioxide nanoparticles in mice fetoplacental unit. J Biomed Mater Res A 106(2): 580-589.
-
Li L, Liu T, Fu C, Tan L, Meng X, et al. (2015) Biodistribution, excretion, and toxicity of mesoporous silica nanoparticles after oral administration depend on their shape. Nanomedicine 11(8): 1915-1924.
-
Lee J, Yu WJ, Song J, Sung C, Jeong EJ, et al. (2016) Developmental toxicity of intravenously injected zinc oxide nanoparticles in rats. Archives of pharmacal research 39(12): 1682-1692.
-
Panzner EA, Jansons VK (1979) Control of in vitro cytotoxicity of positively charged liposomes. J Cancer Res Clin Oncol 95(1): 29-37.
-
Shin DH, Tam YT, Kwon GS (2016) Polymeric micelle nanocarriers in cancer research. Front Chem Sci Eng 10: 348-359.
-
Mendes PL, Pan J, Torchilin V (2017) Dendrimers as nanocarriers for nucleic acid and drug delivery in cancer therapy. Molecules 22(9): 1401.
-
Brown JS, Zeman KL, Bennett WD (2002) Ultrafine particle deposition and clearance in the healthy and obstructed lung. Am J Resp Crit Care Med 166(9): 1240- 1247.
-
Shvedova AA, Kisin ER, Mercer R, Murray AR, Johnson VJ, et al. (2005) Unusual inflammatory and fibrogenic pulmonary responses to singlewalled carbon nanotubes in mice. Am J Physiol 289(5): L698-L708.
-
Risom L, Møller P, Loft S (2005) Oxidative stress-induced DNA damage by particulate air pollution. Mutation Res 592(1-2): 119-137.
-
Wu J, Wang C, Sun J, Xue Y (2011) Neurotoxicity of silica nanoparticles: brain localization and dopaminergic neurons damage pathways. ACS Nano 5(6): 4476-4489.
-
Long H, Shi T, Borm PJ, Maatta J, Husgafvel-Pursiainen K, et al. (2004) ROSmediated TNF-α and MIP-2 gene expression in alveolar macrophages exposed to pine dust. Part Fibre Toxicol 1(1): 3.
-
Li N, Sioutas C, Cho A, Schmitz D, Misra C, et al. (2003) Ultrafine particulate pollutants induce oxidative stress and mitochondrial damage. Environ Health Persp 111(4): 455-460.
-
Xia T, Kovochich M, Brant J, Hotze M, Sempf J, et al. (2006) Comparison of the abilities of ambient and manufactured nanoparticles to induce cellular toxicity according to an oxidative stress paradigm. Nano Lett 6(8): 1794-1807.
-
Xia T, Li N, Nel AE (2009) Potential health impact of nanoparticles. Ann Rev Pub Health 30: 137-150.
-
Sayes CM, Fortner JD, Guo W, Lyon D, Boyd AM, et al. (2004) The differential cytotoxicity of watersoluble fullerenes. Nano Lett 4(10): 1881-1887.
-
Shimizu M, Tainaka H, Oba T, Mizuo K, Umezawa M, et al. (2009) Maternal exposure to nanoparticulate titanium dioxide during the prenatal period alters gene expression related to brain development in the mouse. Part Fibre Toxicol 6: 20.
-
Smith JA, Park S, Krause JS, Banik NL (2013) Oxidative stress, DNA damage, and the telomeric complex as therapeutic targets in acute neurodegeneration. Neurochem Int 62: 764-775.
-
Block M, Wu X, Pei Z, Li G, Wang T, et al. (2004) Nanometer size diesel exhaust particles are selectively toxic to dopaminergic neurons: the role of microglia, phagocytosis, and NADPH oxidase. FASEB J 18(13): 1618-1620.
-
Xia T, Li N, Nel AE (2009) Potential health impact of nanoparticles. An Rev Pub Health 30: 137-150.
-
Peters A (2005) Particulate matter and heart disease: evidence from epidemiological studies. Toxicol Appl Pharm 207(2): 477-482.
-
Geiser M (2010) Update on macrophage clearance of inhaled micro-and nanoparticles. J Aero Med 23(4): 207- 217.
-
Garcia-Garcia E, Andrieux K, Gil S, Kim HR, Le Doan T, et al. (2005) A methodology to study intracellular distribution of nanoparticles in brain endothelial cells. Int J Pharm 298: 310-314.
-
Li N, Sioutas C, Cho A, Schmitz D, Misra C, et al. (2003) Ultrafine particulate pollutants induce oxidative stress and mitochondrial damage. Environ Health Persp 111(4): 455-460.
-
Stefani D, Wardman D, Lambert T (2005) The implosion of the Calgary General Hospital: ambient air quality issues. J Air Waste Manage 55(1): 52-59.
-
Chithrani BD, Ghazani AA, Chan WC (2006) Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells. Nano Lett 6(4): 662-668.
-
Stern ST, Adiseshaiah PP, Crist RM (2012) Autophagy and lysosomal dysfunction as emerging mechanisms of nanomaterial toxicity. Part Fibre Toxicol 9: 20.
-
Wang F, Bexiga MG, Anguissola S, Boya P, Simpson JC, et al. (2013a) Time resolved study of cell death mechanisms induced by amine-modified polystyrene nanoparticles. Nanoscale 5(22): 10868-10876.
-
Xue Y, Wu J, Sun J (2012) Four types of inorganic nanoparticles stimulate the inflammatory reaction in brain microglia and damage neurons in vitro. Toxicol Lett 214(2): 91-98.
-
Roy R, Singh SK, Chauhan L, Das M, Tripathi A, et al. (2014) Zinc oxide nanoparticles induce apoptosis by enhancement of autophagy via PI3K/Akt/ mTOR inhibition. Toxicol Lett 227(1): 29-40.
-
Oberdorster E (2004) Manufactured nanomaterials (fullerenes, C60) induce oxidative stress in the brain of juvenile largemouth bass. Environ Health Perspect 112(10): 1058.
-
Long H, Shi T, Borm PJ, Maatta J, Husgafvel-Pursiainen K, et al. (2004) ROSmediated TNF-α and MIP-2 gene expression in alveolar macrophages exposed to pine dust. Part Fibre Toxicol 1: 3.
-
Brown DM, Donaldson K, Borm PJ, Schins R, Dehnhardt M, et al. (2004) Calcium and ROS-mediated activation of transcription factors and TNF-α cytokine gene expression in macrophages exposed to ultrafine particles. Am J Physiol 286(2): L344-L353.
-
Broday L, Peng W, Kuo MH, Salnikow K, Zoroddu M, et al. (2000) Nickel compounds are novel inhibitors of histone H4 acetylation. Cancer Res 60(2): 238-241.
-
Cangul H, Broday L, Salnikow K, Sutherland J, Peng W, et al. (2002) Molecular mechanisms of nickel carcinogenesis. Toxicol Lett 127(1-3): 69-75.
-
Giri NC, Passantino L, Sun H, Zoroddu MA, Costa M, et al. (2013) Structural investigations of the nickel- induced inhibition of truncated constructs of the JMJD2 family of histone demethylases using X-ray absorption spectroscopy. Biochemistry 52(24): 4168-4183.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells