How do Heterocyclic Amines, Amino Acid Pyrolysates, Inhibit DNA Repair
Heterocyclic amines (HCAs) and β-carbolines, carcinogenic substances, are produced when amino acids and creatine in food react in a high-temperature environment. Some of them are reported to be rodent carcinogens and potent mutagens and to have co- clastogenic effects by inhibiting the incision step in DNA excision repair. In this study, we studied the mechanisms of their inhibition of DNA excision repair. HCAs (Trp-P- 1, IQ, and Me IQ) suppressed comet responses by UV. Since they were antagonistic to novobiocin that is an inhibitor of ATPase subunit of topoisomerase acting the in the incision step of excision repair and the removal of cyclobutane pyrimidine dimers (CPDs) induced by UV was suppressed by Trp-P-1 and IQ, they were considered to inhibit the incision step of excision repair. On the contrary, β-carbolines (harman and norharman) enhanced comet responses by UV. Since they were antagonistic to cytosine-1-β-D arabinofuranoside that is an inhibitor of DNA polymerase acting in the re-synthesis step of excision repair, they were considered to inhibit the re-synthesis step of excision repair. Based on thesis results, HCAs and β-carbolines were concluded to inhibit the incision and re-synthesis steps of excision repair, respectively.
Introduction
Heterocyclic amines (HCAs), carcinogenic substances, are produced when amino acids and creatine in food react in a high-temperature environment. They are especially present in large amounts in burnt parts of fish and meat, and in smoke [1]. About 20 types have now been reported, and have been recognized as carcinogenic by the International Agency for Research on Cancer (IARC) [2]. β-Carbolines, such as harman and norharman, have also been detected in pyrolysates of tryptophan [1]. HCAs have also been reported to be rodent carcinogens [3, 4], and IQ was demonstrated to induce hepatocellular carcinoma in monkeys [5]. We have already shown that HCAs and β-carbolines show co- clastogenic effects by inhibiting the incision step in the nucleotide excision repair (NER) [6]. In this study, we studied the mechanisms of their inhibition of DNA excision repair.
Materials and Methods
Cells
TK+/- heterozygotes of WTK1 human lymphoblastoid cells exhibiting mutant-type p53 (kindly provided by Dr. Honma, National Institute of Health Sciences, Tokyo) were used. WTK1 cells were maintained using RPMI 1640 medium (Nissui Pharmaceutical Co., Ltd.) supplemented with 10% horse serum (SAFC Biosciences), 200 µg/mL sodium pyruvate and 200 µg/mL streptomycin at 37℃ under a 5% CO2 atmosphere.
Cell Treatment with Mutagens: This study was conducted as the experimental design described in our previously study [7, 8]. Cells at 5 x 105 cells/mL were exposed to 40 µg/mL methyl methanesulfonate (MMS, Sigma Chemicals Inc.) or 5 µg/ mL bleomycin (BLM, Wako Pure Chemical Industries, Ltd). Immediately after the chemical treatment, cells were exposed to each HCA with and without DNA repair inhibotors. To irradiate to UVC, 1 mL of cell suspension in saline (5 x 105 cells/mL) in a 6-cm dish was irradiated with a germicide lamp (National GL15, 15 W, Matsushita Electric Industrial Co., Japan) at 10 J/m2.
Detection of CPD: Cells exposed to UVC at 10 J/m2 were post-treated with Trp-P-1, IQ, MeIQ, or harman at 50 µg/mL for 2 h. UVC induced cyclobutane pyrimidine dimers (CPDs) were detected using High Sensitivity CPD ELISA kit (Cosmo Bio Co.,Ltd) immediately after the irradiation and the post- treatment for 2 h with HCAs.
Comet Assay: Cells exposed to UVC were post-treated with each HCAs for 2 h in the presence and absence of the DNA repair inhibitors cytosine-1-β-D-arabinofuranoside (araC, 1.8 mM) with and without novobiocin sodium (NOV, 285.5 µg/mL). Both inhibitors (Wako Pure Chemical Industries, Ltd.) were dissolved in DMSO. Treated cells were suspended in 1% agarose-LGT at 5×105 cells/75 µL, and 75 µL of cell suspension was immediately deposited on a fully frosted slide (Matsunami Glass Ind., Ltd., Osaka, Japan), which was coated with 1% agarose GP-42 and then covered with another slide glass. The slides were placed so as to allow the agarose to gel. The samples on the slides were then immediately exposed to a lysing solution (pH 10) of 2.5 M NaCl, 100 mM EDTA disodium (Na2EDTA), 10 mM Trizma, 1% sarkosyl, 10% DMSO and 1% Triton X-100, and left at 4℃ for 1 h. The slides were then placed on a horizontal gel electrophoresis platform and covered with pH>13 alkaline solution composed of 300 mM NaOH and 1 mM Na2EDTA. The slides were left in solution at 0℃ for 20 min to allow unwinding of the DNA and expression of alkali-labile sites to occur. The power supply was set at 25V and 250 mA. The DNA was subjected to electrophoresis at 0℃ for 20 min and the slides were rinsed with 400 mM Trizma (pH 7.5) to neutralize the excess alkalinity. Each slide was stained with 50 µL of 20 µg/mL ethidium bromide (Wako Pure Chemical Industries, Ltd.) and covered with a cover slip. Fifty cells on one slide per dose (one slide was prepared for each dose) were examined and photographed (black and white ASA 400 Fuji film) at 200× magnification using a fluorescence microscope (Olympus) equipped with a G filter. The tail length of the comet was measured. The effects of chemical treatment on tail length and CPD were analyzed using ANOVA and the Dunnett test.
Results
The effects of HCAs on the tail length induced by UV are shown in Figure 1. In WTK1 cells exposed to UV at 10 J/m2, Trp-P-1, IQ, and MeIQ decreased tail length statistical significantly in the presence and absence of araC. However, they did not affect tail length in the presence of NOV. PhIP did not affect tail length in the presence and absence of repair inhibitors. They did not induce tail length in UV- unirradiated cells. As shown in Figure 2, Trp-P-1, IQ, and MeIQ but not PhIP also decreased tail length statistical significantly in cells exposed to 40 µg/mL MMS but not 5 µg/mL BLM. On the contrary, harman increased tail length induced by UV and MMS but not BLM statistical significantly without araC but not with araC (Figures 3 & 4). Although norharman also increased tail length induced by UV, its enhancing effect was smaller than that of harman and enhancing effect was not observed against MMS. Harman and norharman did not induce tail length in UV-unirradiated cells. CPD induced by UV decreased statistical significantly at 2 h after UV exposure, in the absence of HCAs. The decrease in CPD was also detected in the presence of harman. In the presence of Trp-P-1 and IQ, however, CPD did not decrease statistical significantly at 2 h after UV exposure (Figure 5).
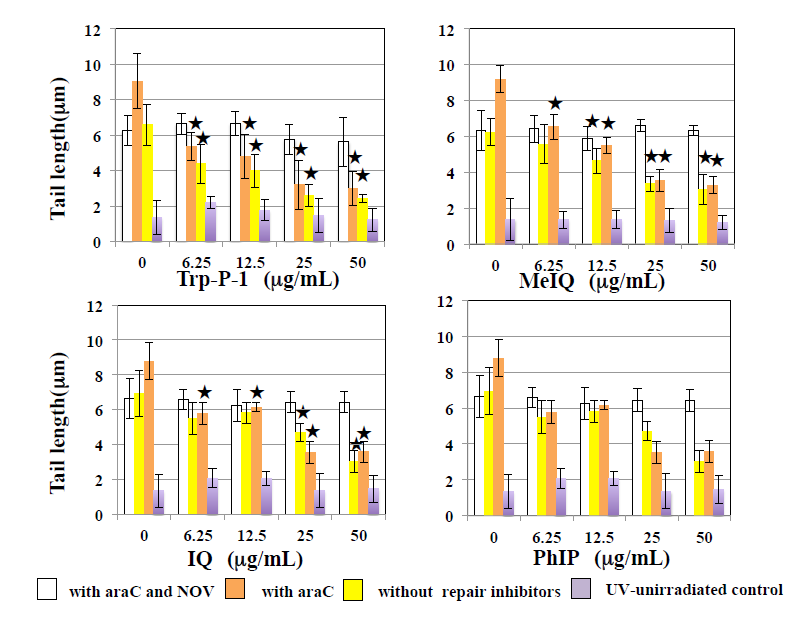
Figure 1: Effects of HCAs on comet responses by UV in WK1 cells. Comet slides were prepared after UV irradiation at 10 J/ m2 followed by the culture for 2 h with HCAs in the presence and absence of 1.8 mM araC, and with and without 285.5 µg/mL NOV. The error bars indicate standard deviation of the mean of three independent trials. Significantly lower than 0 µg/mL HCA control: p<0.05.
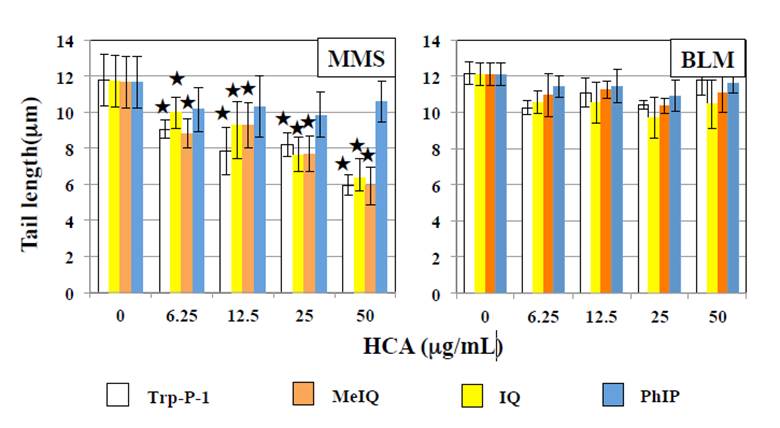
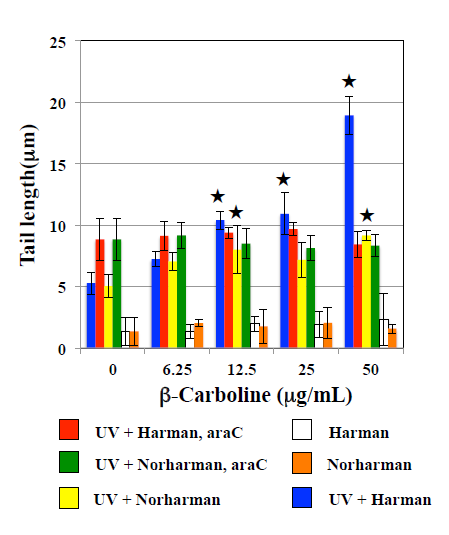
Figure 3: Effects of β-carbolines on comet responses by UV in WK1 cells. Comet slides were prepared after UV irradiation at 10 J/m2 followed by the culture with β-carbolines for 2 h with and with and without 1.8 mM araC. The error bars indicate standard deviation of the mean of three independent trials. ★Significantly higher than 0 mg/mL β-carboline control: p<0.05.
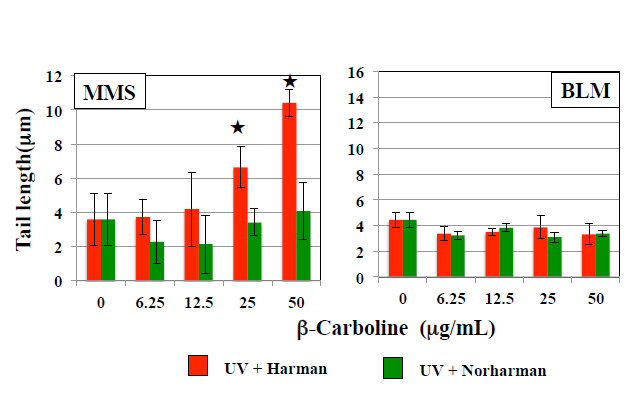
Figure 4: Effects of β-carbolines on comet responses by MMS and BLM in WK1 cells. Comet slides were prepared after culture for 2 h with β-carbolines after 1-h exposure to 40 µg/mL MMS or 5 µg/mL BLM. The error bars indicate standard deviation of the mean of three independent trials. ★ Significantly higher than 0 µg/mL β-carboline control: p<0.05.
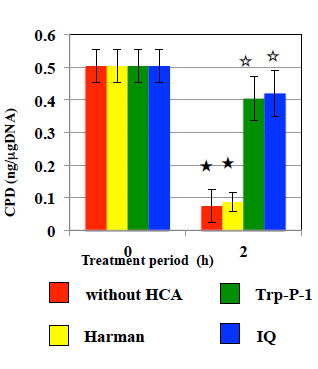
Figure 5: Effects of HCAs and Harman on CPD induced by UV in WK1 cells. CPDs were detected immediately after UV irradiation at 10 J/m2 and after the culture for 2 h with HCAs and Harman following to UV irradiation. The error bars indicate standard deviation of the mean of three independent trials. ★ Significantly lower than HCA 0-h treatment control: p<0.05. ☆Significantly higher than HCA-free control at 2 h: p<0.05.
Discussion
The comet assay detects single strand breaks (SSBs) produced as initial lesions and also those that are generated during the repair of initial lesions such as alkylated bases, bulky base adducts and CPDs [9].
It is known that CPDs are repaired by NER, which consists of the following four steps: recognition of the DNA lesion, excision of a 24–32 nucleotide stretch containing the lesion by dual incision of the damaged DNA strand on both sides, filling in of the resulting gap by DNA polymerase and ligation of the nick.
During the process, SSBs are produced as intermediates, which can be visualized as a comet tail in the comet assay. Therefore, the inhibition of SSB formation in the incision step and promotion of SSB disappearance in the re-synthesis step would result in the suppression of comet responses [7, 8]. The DNA re-synthesis inhibition by araC is due to either direct inhibition of DNA polymerase when araCTP is bound to the dCTP binding site of the enzyme, or indirect inhibition through araCMP incorporation into a repaired region of DNA rendering it unsuitable for further polymerase action [10]. In the case of harman and norharman, their effects on comet responses were canceled by araC, suggesting that araC is antagonistic to them. Therefore, the enhancement of comet responses by them would reflect the increase in comet- detectable SSB by inhibiting the re-synthesis step. HCAs and β-carbolines studied here did not affect tail length in cells not irradiated to UV, showing that they did not induce comet tails by themselves. In the case of Trp-P-1, IQ, and MeIQ, their suppression effects on comet responses were canceled by NOV, suggesting that NOV is antagonistic to them. NOV which is an inhibitor of ATPase subunit of topoisomerase is considered to inhibit the incision step [11] to decrease tail length. UV induced CPD decreased 2 h after the irradiation of UV, showing removal of CPD by NER. This decrease was not observed in the presence of Trp-P-1 and IQ that showed decreases in tail length, supporting the possibility that the decrease in tail length is due to the inhibition of the removal of CPD in NER. HCAs studied affected tail length induced by UV and MMS but not BLM. UV and MMS induce damaged bases that are removed by the NER and base excision repair (BER), respectively, but BLM induces SSBs that are not repaired by the excision repair [12, 13]. These results are considered to coincide with our consideration that HCAs inhibit the excision repair step. Based on those results, HCAs and β-carbolines were concluded to inhibit the incision and re-synthesis steps of excision repair, respectively.
Acknowledgements
This research was conducted in Material and Biological Engineering Course, National Institute of Technology, Hachinohe College as part of a graduation research project by Runa Kobayashi and Azusa Sekishita under the research guidance of course and staffs based on allocation of school educational expenses. The authors acknowledge the Material and Biological Engineering Course, National Institute of Technology, Hachinohe College.
Ethical Approval
It is not applicable.
References
-
Sugimura T, Kawachi T, Nagao M, Yahagi T, Seino Y, et al. (1977) Mutagenic princeple(s) in tryptophan and phenylalanine pyrolysis products Proc Jpn Acad 53(1): 58-61.
-
IARC (1993) Monographs of the Evaluation of the Carcinogenic Risk of Chemicals to Humans. Some Natural Occuring and Synthetic Food Components, Furocoumarins and Ultraviolet Radiation. LARC Scientific Publication, LARC, Lyon, France, 56: 165-195.
-
Sugimura T (1988) Successful use of short-term tests for academic purpose: their use in identification of new environmental carcinogens with possible risk for humans. Mutation Res 205(1-4): 33-39.
-
Sugimura T, Takayama S, Ohgaki H, Wakabayashi K, Nagao M (1990) Mutagens and carcinogens formed by cooking meat and fish: heterocyclic amines. In: Finot PA, et al. (Eds.), The Maillard Nutrition and Physiology, Birkhaeuser Verlag Basel pp: 323-334.
-
Adamson RH, Thorgeirsson UP, Snyderwine EG, Thorgeirsson SS, Reeves JU, et al. (1990) Carcinogenicity of 2-amino-3-methyl-imidazo [4,5- f]quinoline in nonhuman primates: induction of tumors in three macaques J Cancer Res 81(1): 10-14.
-
Sasaki YF, Yamada H, Shimoi K, Kinae N, Tomita I, et al. (1992) Enhancing effects of heterocyclic amines and β-carbolines on the induction of chromosome aberrations in cultured mammalian cells Mutation Res 269(1): 79-95
-
Hayashishita S, Nakamura T, Kikuchi Y, Elizabeth GH, Matsumoto K, et al. (2022) Inhibiting and enhancing effects on DNA repair of rare metal elements in cultured Human lymphoblastoid cells JOCAMR 18(4): 34-41
-
Tahara Y, Hayashishita S, Nakamura T, Murashige R, Yamasaki K, et al. (2022) Genotoxicity-Suppressing Effect of Sophora japonica L. Aqueous Extract. Journal of Complementary and Alternative Medical Research 17(4): 16-26.
-
Fairbairn DW, Olive PL, O Neill KL (1995) The comet assay: a comprehensive review. Mutat Res 339(1): 37- 59.
-
Rojas E, López MC, Valverde M (1999) Single cell gel electrophoresis assay: methodology and applications. J Chromatogr B Biomed Sci Appl 722(1-2): 225-254.
-
Dresler SL, Robinson Hill RM (1987) Direct inhibition of UV- induced DNA excision repair in human cells by novobiocin, coumerycin and nalidixic acid. Carcinogenesis 8(6): 813-817.
-
Mohrenweiser HW, Jones IM (1998) Variation in DNA repair is a factor in cancer susceptibility: a paradigm for the promises and perils of individual and population risk estimation Mutat Res 400(1-2): 15-24.
-
Vig BK, Lewis R (1978) Genetic toxicology of bleomycin. Mutat Res 55(2): 121-145.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells