Impact of Sting Pathway in Inflammation and Covid-19 Pathogenesis
The recent encounter by Sars-Cov2 (COVID-19) presented the need of having stronger immune system as the first line of body’s defense mechanism. Stronger the immune response better will be the pathogen recognition. Better understanding of immune response mechanism and functioning of its components will pave the route for defending against these pathogens. Innate immune defense being non-specific in nature identifies both DNA and RNA viruses as well as other pathogens attacking the body. Presence of foreign nucleic acids bound to the specific pattern recognition receptors (PRRs) which include toll-like receptors (TLRs), cGAS-STING, Nod-like receptors (NLR) and RNA helicase receptors (RIG-1 and MDA5) triggers a signaling cascade. These signaling channels direct to the production of type-1 interferons and other cytokine/chemokine effectors of innate immune system which in turn activate adaptive immune response. STING pathway as a prime innate detector of self and non-self-nucleic acids plays an important role in the development of inflammation followed by type-1 interferon production. COVID-19 being involved with the respiratory failure shows the main characteristic feature of cytokine storm which direct to hyper inflammation. These hyper-inflammatory responses worsen the immune compromised state of the patients which makes this disease as the leading cause of mortality worldwide. Understanding the underline mechanisms involved in and regulating the process of hyper inflammation would provide an effective treatment to reduce the mortalities. Deregulation of STING pathway is involved in various inflammatory diseases that make it a potent target to understand its mechanism of work during viral attack. Many recent studies have further supported the importance of delayed STING signaling in the pathogenesis of COVID-19 during the second phase of this disease. This review mainly focuses on the role of STING in inflammation and its function in our immune system which can be harnessed to tackle the recent pathogeneses of COVID-19.
Introduction
Novel Coronavirus disease commonly known as COVID-19 caused a pandemic with globally 823,626 confirmed (72736) and 40598 deaths(4193) cases in the year 2019 [1]. The spread of this disease hit almost every country with India accounting for 56,342 positive cases in the year 2020 [2]. COVID-19 as an acute respiratory distress syndrome (ARDS) affects mainly the lung tissue. The most common symptoms related to this disease are sputum production, pneumonia, high body temperature, fatigue, lymphopenia (reduced number of CD4+, CD 8+ and NK-cells), cytokine storm and loss of smell and taste [3]. SARS-CoV-2 pathologically shows resemblance with two other coronavirus infections SARS (Severe Acute Respiratory Syndrome) and MERS (Middle East Respiratory Syndrome) [4].
SARS-CoV-2 virus genome show sequence similarity ~98% with RaTG13 bat coronavirus. Bats genomes carry mutation in STING (lack of serine 358 residues) that makes them incapable to detect damaged dsDNA [5]. Bats are considered to be the natural carrier of many viruses recently now thepandemic COVID-19 virus. COVID-19 infected patient’s plasma contains increased concentrations of pro- inflammatory cytokines/chemokines such as IL-6, IL-2, IL- 7, IFN-γ, MCP1 (monocytes chemoattractant protein-1), IL- 10, and MIP1A (macrophage inflammatory protein-1A) that causes local hyper inflammation [6]. This cytokine shock is associated with the disease severity and increased morbidity. Many clinical trials have been focused on the blockade of these cytokines to overcome hyper-inflammatory response. Treatment of COVID-19 patients with tocilizumab (IL-6 inhibitor) improved the condition of patients and decreased the local inflammatory response but the effect was limited [7, 8]. Anti-inflammatory drugs were also used for the treatment of COVID-19 Table 1 that presented the data which confirmed with reduced inflammation, the therapeutic outcome in patients can be improved. The present studies on the scenario of COVID-19 shows that the older people and people with co-morbidities are more susceptible to this hyper- inflammatory response generated [9].
This altered inflammatory response with aging affects the immune response to cope up with this viral infection [10]. But younger childrens are also pre-disposed to this deadly disease. Covid-19 in childrens show better prognosis than adults with elevated inflammatory markers with mild severity but this scenario can be changed in the second wave of this disease [11]. Cytokine storm is the principle cause for the increasing morbidity due this disease. STING an endoplasmic reticulum transmembrane protein acts as an essential DNA sensing mechanism for the production of type- 1 interferons (IFNs) and pro-inflammatory cytokines during viral/microbial entry, auto-inflammatory diseases and cancer [12]. STING also senses other cyclic di-nucleotides (CDNs) in the cytosol such as 2’,3’-cGAMP, 3’,3’-cGAMP, c-di-GMP, c-di- AMP, disrupted DNA and mtDNA for the production of type- 1-interferons [13, 14, 15]. Studies on STING protein encoded by TMEM173 gene validated its role to act as an important sensory link of both cytosolic DNA and RNA in an immune- compromised state [16].
Inflammatory autoimmune diseases such as Aicardi- Goutieres syndrome, systemic lupus erythematosus (SLE), encephalitis, type-1 diabetes are the result of over- activated STING that stimulates increased type-1 interferon production [17, 18, 19]. When it comes to viral proteins they pose a challenge to the anti-viral machinery of the immune system by developing different blocking mechanisms. In an experimental study on six different viral proteins in-vitro and in-vivo it was shown that inhibition of apoptosis machinery causes necroptosis (induced inflammation) through STING and RIG-1 activated pathway [20]. Production of type- 1 interferons provides protective role against pathogen infections and inflammatory diseases. Clinical studie have supported that serum of COVID-19 patients lack sufficient levels of type-I and type-III interferons compared to the other pro-inflammatory cytokines [20]. When we look at the outbreak of two other viruses belonging to family of coronaviruses such as MERS and SARS they show same behavior by restricting the production of type I and type-III interferons compared to the increased production of other inflammatory cytokines [21, 22].
STING activation in response to viruses or virus induced leakage of mtDNA translocates TBK1 and IRF3 from ER (endoplasmic reticulum) to Golgi apparatus for induction of IFNs and ISGs (interferon stimulated genes) [23].To escape the immune response viruses encode different methods to modulate the STING response. COVID-19 patient’s show delayed STING over-activation followed by the DNA damage/ leakage of mtDNA after encounter by SARS-CoV2 virus. This delayed response of STING in COVID-19 correlates with STING gain of function in SAVI syndrome which presents CD4+ and CD8+ lymphocytopenia followed by the over-production of the inflammatory cytokines [24].Thus understanding the underline inflammatory mechanism through the route of STING pathway in viral infections would open new therapeutic method to treat COVID-19 patients.
| Dose | Function | Efficacy | |
|---|---|---|---|
| Glucocorticoids | 1-2mg/kg per day | Suppress cytokine storm | low |
| Tocilizumab | 400mg (intravenously) | IL-6 inhibitor | high |
| Chloroquine and Hydroxychloroquine | 300-500mg 2times/day | Regulate pH dependent viral entry and cell signaling for production of pro-inflammatory cytokines. | high |
| Convalescent plasma therapy | Plasma transfusion | Decreased viral load | median |
| Covaxin | 2-dose | Whole inactivated virion cell | Greater response rate 86.8% in patients with previous virus exposure. |
| Covishield | 2-dose | Weakend adenovirus | 70.4% efficacy |
Table 1: Clinical trial drugs and vaccines tested and available against COVID-19 that showed potent therapeutic role in this dise
Function of Sting Protein in Inflammation
Inflammation is an immune response against wide range of substances which includes autophagic/necrotic molecules, altered self/non-self-nucleic-acids and various other environmental molecules. The process of inflammation involves both innate and adaptive immune system for maintaining the tissue homeostasis [18]. Despite its protective effects the process when uncontrolled might result in various auto-inflammatory diseases, metabolic disorders and cancer. Inflammation also causes trafficking of monocytesin response to MCP-1 stimulus via blood stream that is beneficial for the clearance of invading pathogens [19]. At the inflammatory site these monocytes differentiate into DCs and macrophages that activate CD4+ cells which further trigger the production of cytokines [20]. STING pathway is the predominating signaling mechanism in macrophages during the inflammatory response. Study conducted on genetically modified mice showed that this pathway in macrophages is promoter of inflammation in acute pancreatitis [21]. In another study on human macrophage cell line (THP-1) infected with HIV, HSV-1 and CMV viral genome produced interferon response via cGAS-cGAMP-STING-TBK1 pathway with involvement of cellular co-factor IFI16 [22]. In recent experimental study on silica induced lung inflammation in- vitro and in vivo results confirmed the role of STING pathway to induce lung inflammation upon sensing of damaged DNA particles in cytosol [23] (Figure 1).
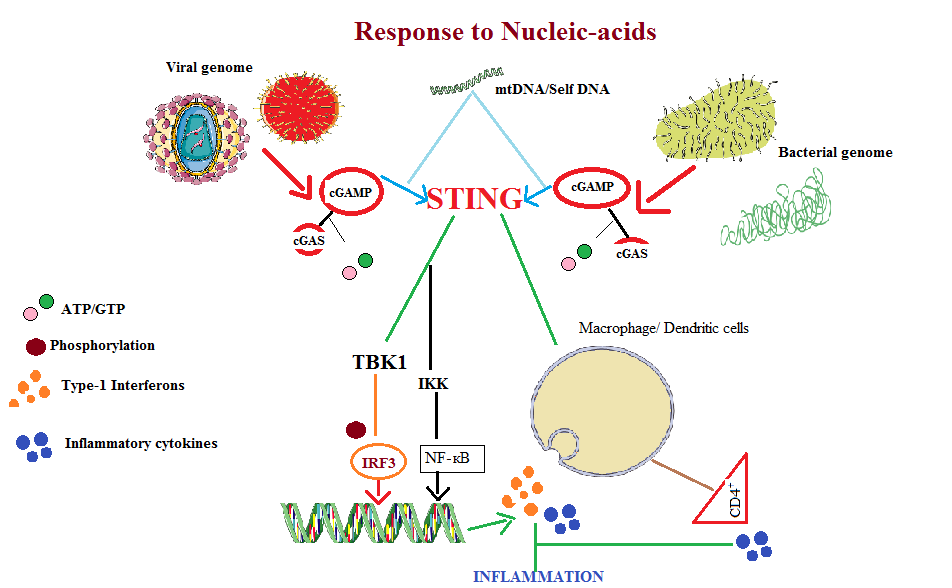
Figure 1: STING pathway induces the production of Type-1 Interferons and pro-inflammatory cytokines in response to the intracellular pathogens or towards the mtDNA (mitochondrial DNA)/Self DNA particles present in the cytosol. Activation process of this pathway then stimulates the downstream TBK1 (TANK-binding kinase1) which phosphorylates IRF3 (Interferon regulator factor3). In nucleus it induces the transcription of genes encoding type-1 Interferons and pro-inflammatory cytokines. In response to RNA STING interacts with RIG1 (Retinoic-acid inducible gene 1) via MAVS (mitochondria antiviral signaling) to induce type-1 interferon production via IкB (IKK) and NF-кB.
Signaling mechanisms RIG-1-MAVS-STING RNA with cGAS-STING DNA sensing in collaboration generate efficient anti-viral responses [24] (Figure 2). During inflammation process the dysregulated STING pathway leads to over- production of type-1 Interferons and pro-inflammatory cytokines leading to cytokine storm (Table 2).
| Nature | Origin | Function | Ref | |
|---|---|---|---|---|
| Rv0753c(MmsA), mitochondrial enzyme | Antagonist of STING | Mycobacterium tuberculosis virulence strain H37Rv. | Modulate type I interferon production by endogenous binding with STING that inhibit STING-TBK1-IRF3 pathway | [18,19] |
| Menadione(2-methyl-1,4-naphthoquinone) | Antagonist of STING | Synthetic form vitamin K | Anti-hemorrhagic, antibacterial, anti-inflammatory, generation of ROS | [20, 21] |
| SU0268 | Antagonist to OGG1 (8-oxoguanine DNA glycosylase-1) and its downstream STING pathway. | Tetrahydroquinoline sulfonamide derivatives. | Anti-inflammatory. | [22-24] |
| H-151 | Antagonist to STING | Selective peptide inhibitor | Anti-inflammatory | [25,26] |
| ISD017 | Antagonist to STING | Peptide inhibitor | Anti-inflammatory | [27] |
| Remdesivir(GS-5734) | Antagonist to STING | Phosphoramidate nucleotide drug | Anti-inflammatory | [28-30] |
| IACS-8803, IACS-8779 | Agonist of STING | 2',3'-thiophosphate CDN analogs | Leads to activation of the STING pathway | [31, 32] |
| Manassantin B | Agonist of STING | Natural product extracted from Saururus chinensis bail. | Increase the production of mitochondrial ROS activates STING-TBK1-IRF3 signalling and reduction of pro-inflammatory cytokines/chemokines. | [33,34] |
| Suramin | Antagonist of cGAS | Polysulfonated naphthylurea anti-parasitic drug | Anti-inflammatory and reverse transcriptase inhibitor | [35-37] |
| Amidobenzimidazole (diABZI) | Agonist of STING | Small synthetic chemical compounds which have high potency than cGAS for STING | Anti-viral and anti-cancer properties | [38, 39] |
Table 2: Table contents showing various agonist and antagonist of STING pathway.
Role of Sting Pathway in Immunity
Any type of biological stress and disease due to entry of microbes and viruses pose threat to the body functioning. These pathogens after entry in the cell cytoplasm release viral/ microbial dsDNA/ssRNA that triggers the immune response. Immune function via immune cells/signaling mechanism and other molecules builds-up the protective environment against any invading pathogens attack. Viruses/bacteria are detected by immune cells via different sensors present inside the cell which include RIG-1 like receptors(RLRs), cyclic GMP-AMP synthase(cGAS), Interferon-γ(IFNγ)-inducible protein16(IFI16), toll like recetors(TLRs), AIM2 [25, 26]. IFI- 16, RIG-1 and cGAS through the common STING route located on the endoplasmic reticulum and golgi-body heightens the innate immune response in the immune cells. Upon activation cGAS leads to formation of the second messenger (2’-3’cGAMP) which acts as the ligand for STING initiating type-1 interferon and pro-inflammatory cytokine signaling [34]. Encounter of RNA viruses inside the cell releases ssRNA/dsRNA; they also make the accumulation of RNA- DNA hybrids that trigger TLR-9, cGAS and RIG-1 combined sensor response [35]. CGAS apart from nucleic-acid sensor also control various other cellular processes such as inflammation, energy homeostasis, muscular degeneration, autophagy, mitophagy and macrophage transformation [36]. The biological link between the viral entry and its pathogenesis is through the replication of its viral nucleic-acid particles. In an experimental study on HSV-1 γ134.5(Herpes Simlpex virus-1) protein binding with STING in turn inhibits its phosphorylation that silence the STING pathway leading to its pathogensis [37]. Role of STING in sensing dsDNA being its main function, in-vivo study on murine fibroblast cells have shown its alternative role by which it restrict the replication of RNA viruses compared to the STING deficient cells which are incapable [38]. Another in-vivo study on enveloped RNA viruse (Influeza A) have shown an increase in type-1 interferon production by their binding and inhibiting STING dimerization via fusogenic liposomes independent of the main route which involve cGAS [39]. As for its auto- inflammatory role over activation or mutations in the STING protein lead to increased production of type-1 interferons (type-1 interferonopathies) and pro-inflammatory cytokines which cause auto-inflammatory diseases such as pancreatitis, aicardi-Goutieres syndrome(AGS), STING associated vasculopathy with onset in infancy(SAVI), systemic lupus erythematosus(SLE) and non-alcoholic fatty liver to non- alcoholic steatohepatitis (NASH) (Table 3) [40, 41, 42].
| DNA/RNA Sensor | Gene Encoded | Structure | Immune Function |
|---|---|---|---|
| IFI16 | Interferon gamma inducible protein 16 gene encoded on chromosome 1. | Member of HIN-200 protein family containing two HIN domains and one PYRIN domain. Located in cytoplasm and nucleus. | Senses both viral and bacterial single/ double/quadruplex stranded DNA particles that stimulates innate immune response for DNA damage and cell proliferation by formation of inflammasomes. |
| RIG-1 | Retinoic acid inducible gene 1 encoded by gene DDX58 present on chromosome 9. | Protein has central helical structure and carboxyl terminal domain localized in cytolplasm with ATP dependent repressor domain on C-terminus which binds to RNA molecule and two caspase activation and recruitment domains (CARDs) on N-terminus which interact with mitochondrial antiviral signaling protein(MAVS) | RIG-1 is activated by binding to RNA molecule containing triphosphate (PPP) or uncapped diphosphate (PP) or unmethylated at its 2’-O position of their 5’-end. Activates innate immune response against autoimmunity and viral response. |
| AIM2 | Interferon inducible protein or Absent in melanoma 2 is encoded by gene AIM 2 present on chromosome 1. | AIM2 protein structure contain DAPIN or PYRIN signaling domain on N-terminal and HIN-200 DNA binding domain on C-terminal | Senses dsDNA during viral or microbrial infection also in inflammatory diseases by the formation of inflammasome. |
| cGAS | Cyclic GMP-AMP synthase protein is encoded by cGAS gene located on chromosome. | Cyclic GMP-AMP is a cytosolic protein binding to STING on activation. Structure of human cGAS consist of N-terminal helical extension with C-terminal domain and unique zinc- ribbon domain | cGAS upon activation by pathogenic dsDNA leads to Type I interferon response via STING. cGAS-STING pathway also play role in the cancer immunity. |
| PKR | Protein Kinase R protein is encoded by EIF2AK2 gene located on chromosome 2. | Structurally PKR consists of N-terminal domain consists of two RNA binding motifs (dsRBM1 ,dsRBM2) and linker connecting to C-terminal domain. | Function in regulation of cellular stress sensing pathways and in recognition of dsRNA from viruses. |
| TLR9 | Toll like receptor9 protein is encoded by TLR9 gene present of chromosome 3. | TLR 9 protein structure located on endoplasmic reticulum, endosomes and lysosomes contain N-terminal ligand binding extracellular domain with leucine rich repeats(LRR) connected via helical transmembrane domain with C-terminal intercellular Toll IL-1 domain(TIR). | TLR-9 function in sensing of pathogenic dsDNA molecules which initiates cytokine release that mediated immune response. It also function in other cellular process like cell proliferation, stress, invasion and cancer progression. |
Table 3: Table contents lists some of the important nucleic-acid sensors present inside the human cell [28- 32].
Sting Pathway Regulation
A range of biological molecules have shown to regulate the STING pathway which eventually alters the immune response in an anti-viral state. Many of which being the positive regulators that increase the function of STING for production of IFNs in anti-viral state and other being negative regulators that check its activity to prevent auto- inflammatory response. The cGAS-STING pathway acts as one of the activating signaling mechanism in macrophages. Many biological molecules regulating STING by their binding with the targets in cGAS-STING pathway influence macrophages as shown by the experimental study on microRNAs (miR29a/ miR378b) and TMEM203being as positive regulators [43, 44]. STING pathway key functional role is to induce the production of Type-1 interferons (IFN-1) in an anti-viral environment upon activation by pathogenic dsDNA. STAT-1 acts as the positive regulator for the production of IFN-1 via STING with its two binding sites on the STING promoter site [45].Post- translational modification of STING protein require lysine K-27/ K63 linked E3 ubiquitination for its functional activity, USP13 (Ubiquitin-specific protease 13) deconjucated with K-27 and TRIM30α in an in-vivo experiments shown to be the negative regulators of STING activity [46, 47]. STIM1 an endoplasmic reticulum withholding STING protein prevents its over-activity which otherwise cause autoimmunity and auto-inflammatory conditions [48].Viruses evolve different mechanisms to escape the immune response one of which involves by the inhibition of STING pathway as shown by papain-like proteases (PLP) in human coronovirus (HCoV) NL63/severe acute respiratory syndrome (SARS) infections or by the degradation of β-arrestin-2 protein an activator of cGAS-STING pathway [49, 50].There is diversity in the regulatory mechanisms controlling the action of STING pathway; additional studies are ongoing for further understanding its regulation.
Sting Relation with Covid-19
Irregulated function of STING pathway plays a fundamental role in the disease severity. Expression of STING is mostly seen in lung alveolar epithelial cells, spleen cells, immune cells (macrophages, B-cells, T-cells and NK cells) and endothelial cells. Over-expression of STING is validated to cause cytokine storm in mice when administrated with STING agonist DMXAA (40mg) and death compared to STING knockout mice or DMXAA (10mg) [51]. TMEM173 STING encoded gene is known to show heterogeneity among human population [52]. SAVI (STING-associated vasculopathy with onset in infancy) is an example of dominant activating mutation in TMEM173 that show similar symptoms with COVID-19 pathogenesis such as pulmonary inflammation of lung, increased IFN-release, fever and inflammatory vasculopathy. In the second phase of COVID-19 pathogenesis, delayed STING activation due to self-damaged/mt-DNA leads to IFN-βsecretion as seen in murine models of SARS due to excess influx of inflammatory macrophages that would worsen the disease [53]. ACE2 receptor binding with SARS- CoV2 releases angiotensin II signaling that activates STING validated from the results on mice models. This over-activated STING can lead to thrombotic coagulopathy, endothelial inflammation, and increased synthesis of (IFN-β IFN T β-6 and IL-1β) pro-inflammatory cytokines [54]. COVID-19 cases in childrens explained on basis of delayed enhanced signaling of STING in immune cells (macrophages, dendritic cells and B-cells) lead to triggering of CD4+, CD8+ cells and NK-cells that advance SARS-CoV2 or even cause late recurrence with severe fatality [55]. Targeting cGAS, ALK and STING to bring down the cytokine storm and hyper-inflammation in COVID-19 patients for which 6-FDA approved drugs were screened (sorafenib, entecavir, penciclovir, felbamate, genistein and eltrombopag) in which ALK (anaplastic lymphoma kinase) inhibitors showed positive result in blocking the STING dependent inflammatory response [56]. Many agonist and antagonist compounds have been studied mainly that alter the functioning of STING showed promising results in anti-viral, anti-inflammatory and anti-tumor activities. These results pointed STING pathway as potent target to be considered for regulating the cytokine storm in COVID-19 patients which could worsen in the second and third phase of this disease.
Conclusion
The role of STING pathway as a functional anti-viral response is well studied. STING is known for the sensing of cyclic di-nucleotides (CDNs), disrupted DNA particles and mtDNA release from the infected cells during the pathogenic response. RIG-I as a viral RNA cytosolic sensor inside the cells collaborates with the STING protein which eventually activates mitochondrial receptor protein MAVS (Mitochondrial antiviral signaling protein). In an antiviral state activated STING cause release of IFNs which eventually activates innate and adaptive immune response. IFNs as an anti-viral response protect the neighboring cells from the further damage by the viral attack. COVID-19 as an acute respiratory syndrome accounts highest mortality worldwide in part of its efficient viral replication, immune suppression and cytokine storm. The prolonged SARS-CoV2 pathogenesis eventually damages the lung cells which cause release of the disrupted self-DNA and mtDNA into the cytosol. STING pathway activated in response of these particles is beneficial when function in a controlled regulated mechanism. From the experimental clinical studies on COVID-19 patients in relation to the healthy controls have showed decreased concentration of IFNs compared to high concentration of pro-inflammatory cytokines in their plasma. This altered functioning of STING is explained in the literature either due to the polymorphic nature of the TMEM173 gene or the delayed over expression of this protein both of which leads to hyper-inflammatory state of condition. STING as a main sensory protein is also expressed in hematopoietic, monocytes, T-lymphocytes and NK-cells. COVID-19 patients also presented lymphopenia condition with markedly expanded population of monocytes which further add to this cytokine storm via STING activated pathway. Many studies on mice models presented the data which showed the reduced cytokine production by blocking the STING activated pathway. Also the drugs tested against COVID-19 show effective results when target to dampen the cytokine storm. Thus STING pathway as the route of cytokine storm could be the beneficial target in COVID-19 management. Better understanding of its delayed over-activation and the components involved in its signaling mechanism could prove to be the better therapeutic option in COVID-19 pathogensis which will over-come the hyper-inflammatory condition .
References
-
WHO (2020) Coronavirus disease 2019 (COVID-19): situation report, pp: 72.
-
Kumar SU, Kumar DT, Christopher BP, Doss C (2020) The rise and impact of COVID-19 in India. Frontiers in medicine 7: 250.
-
Rothan HA, Byrareddy SN (2020) The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. Journal of autoimmunity 109: 102433.
-
Xu Z, Shi L, Wang Y, Zhang J, Huang L, et al. (2020) Pathological findings of COVID-19 associated with acute respiratory distress syndrome. The Lancet respiratory medicine 8(4): 420-422.
-
Mann DL (2020) SARS-CoV-2 and Bats: From Flight to Fighting COVID-19. J Am Coll Cardiol Basic Trans Science 5(5): 545-546.
-
Tay MZ, Poh CM, Rénia L, MacAry PA, Ng LF, et al. (2020) The trinity of COVID-19: immunity, inflammation and intervention. Nature Reviews Immunology 20: 363-374.
-
Xu X, Han M, Li T, Sun W, Wang D, et al. (2020) Effective treatment of severe COVID-19 patients with tocilizumab. Proceedings of the National Academy of Sciences 117(20): 10970-10975.
-
Luo P, Liu Y, Qiu L, Liu X, Liu D, et al. (2020) Tocilizumab treatment in COVID‐19: A single center experience. Journal of medical virology 92(7): 814-818.
-
Nikolich-Zugich J, Knox KS, Rios CT, Natt B, Bhattacharya D, et al. (2020) SARS-CoV-2 and COVID-19 in older adults: what we may expect regarding pathogenesis, immune responses, and outcomes. Geroscience 42(2): 505-514.
-
Goldstein DR (2010) Aging, imbalanced inflammation and viral infection. Virulence 1(4): 295-298.
-
Ciuca IM (2020) COVID-19 in children: an ample review. Risk Manag Healthc Policy 13: 661-669.
-
Barber GN (2015) STING: infection, inflammation and cancer. Nature Reviews Immunology 15(12): 760-770.
-
Cai X, Chiu YH, Chen ZJ (2014) The cGAS-cGAMP- STING pathway of cytosolic DNA sensing and signaling. Molecular cell 54(2): 289-296.
-
Riley JS, Tait SW (2020) Mitochondrial DNA in inflammation and immunity. EMBO reports 21(4): e49799.
-
Burdette DL, Vance RE (2013) STING and the innate immune response to nucleic acids in the cytosol. Nature immunology 14(1): 19-26.
-
Ahn J, Gutman D, Saijo S, Barber GN (2012) STING manifests self DNA-dependent inflammatory disease. Proceedings of the National Academy of Sciences 109(47): 19386-19391.
-
Mohamed E, Lemos H, Huang L, Chandler P, Souza G, et al. (2014) Activation of the stimulator of interferon genes (STING) adaptor attenuates experimental autoimmune encephalitis. J Immunol 192(12): 5571-5578.
-
Ahn J, Ruiz P, Barber GN (2014) Intrinsic self-DNA triggers inflammatory disease dependent on STING. The Journal of Immunology 193(9): 4634-4642.
-
Schock SN, Chandra NV, Sun Y, Irie T, Kitagawa Y, et al. (2017) Induction of necroptotic cell death by viral activation of the RIG-I or STING pathway. Cell Death & Differentiation 24(4): 615-625.
-
Zhang W, Zhao Y, Zhang F, Wang Q, Li T, et al. (2020) The use of anti-inflammatory drugs in the treatment of people with severe coronavirus disease 2019 (COVID-19): The experience of clinical immunologists from China. Clinical Immunology 215: 108393.
-
Gao J, Tian Z, Yang X (2020) Breakthrough: Chloroquine phosphate has shown apparent efficacy in treatment of COVID-19 associated pneumonia in clinical studies. Bioscience trends 14(1): 72-73.
-
Chen Z, Hu J, Zhang Z, Jiang S, Han S, et al. (2020) Efficacy of hydroxychloroquine in patients with COVID-19: results of a randomized clinical trial. MedRxiv.
-
Tobaiqy M, Qashqary M, Al-Dahery S, Mujallad A, Hershan AA, et al. (2020) Therapeutic Management of COVID-19 Patients: A systematic review. Infection Prevention in Practice 2(3): 100061.
-
Henson PM (2005) Dampening inflammation. Nature immunology 6(12): 1179-1181.
-
Shi C, Pamer EG (2011) Monocyte recruitment during infection and inflammation. Nature reviews immunology 11(11): 762-774.
-
Iijima N, Mattei LM, Iwasaki A (2011) Recruited inflammatory monocytes stimulate antiviral Th1 immunity in infected tissue. Proc Natl Acad Sci 108(1): 284-289.
-
Zhao Q, Wei Y, Pandol SJ, Li L, Habtezion A, et al. (2018) STING signaling promotes inflammation in experimental acute pancreatitis. Gastroenterology 154(6): 1822-1835.
-
Jønsson KL, Laustsen A, Krapp C, Skipper KA, Thavachelvam K, et al. (2017) IFI16 is required for DNA sensing in human macrophages by promoting production and function of cGAMP. Nature communications 8(1): 1-7.
-
Benmerzoug S, Rose S, Bounab B, Gosset D, Duneau L, et al. (2018) STING-dependent sensing of self- DNA drives silica-induced lung inflammation. Nature communications 9(1): 1-9.
-
Zevini A, Olagnier D, Hiscott J (2017) Crosstalk between cytoplasmic RIG-I and STING sensing pathways. Trends in immunology 38(3): 194-205.
-
Abe T, Marutani Y, Shoji I (2019) Cytosolic DNA‐sensing immune response and viral infection. Microbiol Immunol 63(2): 51-64.
-
Chan YK, Gack MU (2016) Viral evasion of intracellular DNA and RNA sensing. Nat Rev Microbiol 14(6): 360- 373.
-
Hároníková L, Coufal J, Kejnovská I, Jagelská EB, Fojta M, et al. (2016) IFI16 preferentially binds to DNA with quadruplex structure and enhances DNA quadruplex formation. PLoS One 11(6): e0157156.
-
Rehwinkel J, Gack MU (2020) RIG-I-like receptors: their regulation and roles in RNA sensing. Nat Rev Immunol 20(9): 537-551.
-
Sharma BR, Karki R, Kanneganti TD (2019) Role of AIM2 inflammasome in inflammatory diseases, cancer and infection. Eur J Immunol 49(11): 1998-2011.
-
Kranzusch PJ, Lee AS, Berger JM, Doudna JA (2013) Structure of human cGAS reveals a conserved family of second-messenger enzymes in innate immunity. Cell Rep 3(5): 1362-1368.
-
Gal-Ben-Ari S, Barrera I, Ehrlich M, Rosenblum K (20019) PKR: a kinase to remember. Front Mol Neurosci 11: 480.
-
Martínez-Campos C, Burguete-García AI, Madrid-Marina V (2017) Role of TLR9 in oncogenic virus-produced cancer. Viral Immunol 30(2): 98-105.
-
Bai J, Liu F (2019) The cGAS-cGAMP-STING pathway: a molecular link between immunity and metabolism. Diabetes 68(6): 1099-1108.
-
Li Y, Wilson HL, Kiss-Toth E (2017) Regulating STING in health and disease. J Inflamm (Lond) 14: 11.
-
Christensen MH, Paludan SR (2017) Viral evasion of DNA-stimulated innate immune responses. Cell Mol Immunol 14(1): 4-13.
-
Bai J, Liu F (2019) The cGAS-cGAMP-STING pathway: a molecular link between immunity and metabolism. Diabetes. 68(6):1099-108.
-
Pan S, Liu X, Ma Y, Cao Y, He B, et al. (2018) Herpes simplex virus 1 γ134. 5 protein inhibits STING activation that restricts viral replication. J Virol 92(20): e01015-18.
-
Franz KM, Neidermyer WJ, Tan YJ, Whelan SPJ, Kagan JC, et al. (2018) STING-dependent translation inhibition restricts RNA virus replication. Proc Natl Acad Sci U S A 115(9): E2058-E2067.
-
Holm CK, Rahbek SH, Gad HH, Bak RO, Jakobsen MR, et al. (2016) Influenza A virus targets a cGAS-independent STING pathway that controls enveloped RNA viruses. Nat Commun 7: 10680.
-
Landman SL, Ressing ME, van der Veen AG (2020) Balancing STING in antimicrobial defense and autoinflammation. Cytokine Growth Factor Rev 55: 1-14.
-
Kumar V (2019) A sting to inflammation and autoimmunity. J Leukoc Biol 106(1): 171-185.
-
Liu Y, Jesus AA, Marrero B, Yang D, Ramsey SE, et al. (2014) Activated STING in a vascular and pulmonary syndrome. N Engl J Med 371(6): 507-518.
-
Shah AU, Cao Y, Siddique N, Lin J, Yang Q, et al. (2019) miR29a and miR378b Influence CpG-Stimulated Dendritic Cells and Regulate cGAS/STING Pathway. Vaccines 7(4): 197.
-
Li Y, James SJ, Wyllie DH, Wynne C, Czibula A, et al. (2019) TMEM203 is a binding partner and regulator of STING- mediated inflammatory signaling in macrophages. Proc Natl Acad Sci U S A 116(33): 16479-16488.
-
Ma F, Li B, Yu Y, Iyer SS, Sun M, et al. (2015) Positive feedback regulation of type I interferon by the interferon‐ stimulated gene STING. EMBO Rep 16(2): 202-212.
-
Sun H, Zhang Q, Jing YY, Zhang M, Wang HY, et al. (2017) USP13 negatively regulates antiviral responses by deubiquitinating STING. Nat Commun 8: 15534.
-
Wang Y, Lian Q, Yang B, Yan S, Zhou H, et al. (2015) TRIM30α is a negative-feedback regulator of the intracellular DNA and DNA virus-triggered response by targeting STING. PLoS Pathog 11(6): e1005012.
-
Srikanth S, Woo JS, Wu B, El-Sherbiny YM, Leung J, et al. (2019) The Ca2+ sensor STIM1 regulates the type I interferon response by retaining the signaling adaptor STING at the endoplasmic reticulum. Nat Immunol 20(2): 152-162.
-
Sun L, Xing Y, Chen X, Zheng Y, Yang Y, et al. (2012) Coronavirus papain-like proteases negatively regulate antiviral innate immune response through disruption of STING-mediated signaling. PLoS One 7(2): e30802.
-
Zhang Y, Li M, Li L, Qian G, Wang Y, et al. (2020) β-arrestin 2 as an activator of cGAS-STING signaling and target of viral immune evasion. Nat Commun 11(1): 6000.
- Effects of 5-HTP and Melatonin on the Sleep Cycle of Medical Students
- Adsorption of Bisphenol A on NH4OH- Modified Rice Husk and Sugar Cane Bagasse Biochar
- Comparative Assessment of the Reinforcement Efficiency of Palm Fruit Fibre and Coconut Fibre in High Density Polyethylene (HDPE) Matrix Composite
- Importance of Bio Compounds Naturally Present in Food with Functionality in Animal Metabolism
- Sub-Acute Study on the Cardiotoxic Effects of Monosodium Glutamate Ingestion in Albino Rat
- Weight Management and Its Natural Solutions: A Review