Mechanisms of Immune Dysregulation Induced by Pesticide Exposure and their Implications for Cancer: A Systematic Review
The inappropriate use of pesticides contaminates natural resources and has consequences for human health. Evidence of immune dysregulation caused by pesticides has been presented, but studies linking exposure to these substances with immune dysregulation and its relationship to cancer are scarce. This study systematically reviews the published literature that addresses the topic of immune dysregulation and cancer and its relationship to pesticide exposure. Studies developed in vitro cell culture and in vivo experimental animals and human exposure data were included. The results show that pesticide exposure has a relevant impact on immune dysfunction, negatively affecting immune system cells and cytokine production in innate and adaptive responses. The main consequences reported were suppression of the Th1 response, induction of the Th2 response, cellular balance dysregulation, and cytokine production changes. All reported events are related to the occurrence of immunosuppression. This dysregulation is paramount for cancer progression and may lead to a worse prognosis in chronically exposed populations. These findings also point to the need for further studies, primarily focused on human exposure.
Introduction
The use of pesticides began last century, and despite studies showing that their properties can cause numerous damages to human and animal health, they started to be used as herbicides, insecticides, and fungicides [1, 2]. Furthermore, it is known that the damages caused by these pesticides take into account the route of exposure, the amount absorbed, the frequency of use, the mode of application, the exposure time, and the general health conditions of the exposed individual [3, 4]. Human exposure can occur through various routes, including accidental exposure, through the ingestion of contaminated water and food or inhalation, and occupational exposure, resulting from continuous handling of pesticides during the production process through conventional agriculture [5].
Another important point is in relation to the extensive use of pesticides, responsible for global environmental pollution. After application, their residues reach rivers and lakes through rainwater runoff, causing ecological pollution and affecting the survival of aquatic organisms. In the case of persistent organic pollutants, environmental contamination can occur for decades after their mobilization from contaminated soil [6].
The impacts of pesticide exposure are wide-ranging, making it a public health problem. These chemical agents can accumulate in the body and interact with biochemical processes, disrupting the body’s homeostasis and resulting in alterations of various systems, including immunological disorders [7]. Recent studies suggest that pesticide exposure impairs immunity, reducing defense against microorganisms and altering the production of inflammatory mediators such as cytokines and chemokines, which are critical for immune responses against pathogens, healing processes, and repair, as well as surveillance against tumor development [8, 9].
These inflammatory cytokines play a crucial role in the surveillance response against tumors by directly affecting growth, cell invasion, and metastasis development [10].
However, pesticide exposure hinders the action of this response and consequently creates favorable conditions for the development of neoplasms [11].
Although the identification of chemical compounds that are not naturally produced or found in the human or animal body and are associated with immunotoxicity has grown in importance in recent years, significant gaps remain in understanding the mechanisms by which these compounds disrupt the immune system. This is particularly true regarding pesticide exposure as a risk factor for cancer development in humans [7].
Therefore, this literature review aims to systematize the mechanisms of immune dysregulations associated with pesticide exposure by reviewing in vitro and in vivo studies and their implication in developing neoplasms in exposed populations. To this end, this review comprises data published in the literature from the last 33 years regarding this topic and presents these findings as a systematic review.
Methods
Study Design
This systematic review was based on the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta- Analyses) guidelines [12]. The PICO (Patient, Intervention, Comparison, and Outcome) strategy was used involving the four components: population to be studied (patient); level or type of exposure (intervention); comparison between exposure and non-exposure to pesticides (comparison); effects on the immune system, cells, and cytokines related to the immune system (primary outcome) and cancer (secondary outcome).
Inclusion and Exclusion Criteria
Studies with experimental animals, in vitro, involving cell cultivation, or with human beings that related the topic of exposure to pesticides to the immune system were included in this research. Literature review articles or articles covering any disease other than cancer were excluded from this search. Studies in the form of theses, dissertations, videos or books, letters to the editor or repeated studies were also excluded.
Study Search, Screening, and Selection
The searches were carried out in electronic databases: PubMed and Scientific Electronic Library Online (SciELO). However, results consistent with the established inclusion criteria were not obtained in the SciELO database. A maximum cutoff date of March 1, 2024, was stipulated and no minimum cutoff date was stipulated. The following descriptor terms were used: “pesticides, cytokines, and cancer”; “pesticides, interleukins and cancer”; “pesticides, immunological and carcinogenic”; “pesticides, immune system and cancer”; “pesticides, immunotoxicity and cancer”; “pesticides, immunological alteration and cancer”; “pesticides, immune response, and cancer.” Initially, the studies were classified by title and then by abstract. When the summary was considered relevant, the study was selected for reading in full.
One author performed this screening and any disagreement was discussed with two senior investigators (C.P. and D.M.B.). The reference list of each eligible article was also cross-referenced for additional studies. If two studies involving the same sample reported complementary results that would otherwise be missed if only one was included, both were considered eligible for data extraction. There was no search in the gray literature.
For each observational study included, the following items were extracted: classification by population, year of publication, study design, number of cases and controls, type of exposure, type of pesticide, markers of immunity/ inflammatory dysregulation, mechanisms of immune system dysregulation innate and adaptive, relationship with immune system cells and cytokines involved and clinical outcome (cancer). The collected data were allocated into two groups: I) in vitro and experimental animals; II) humans. Given the heterogeneity of the categories of studies found statistical studies were not used to explore the results found. Data from all included studies are available in Table 1.
Results
In total, the search found 1,272 published studies, of which 27 were selected according to the eligibility criteria above, as detailed in Figure 1. Of the 1,272 studies, 596 were excluded based on reading the title and 603 were excluded after reading of the summary. The full text of the remaining 73 articles was reviewed and ultimately 27 articles were selected for inclusion.
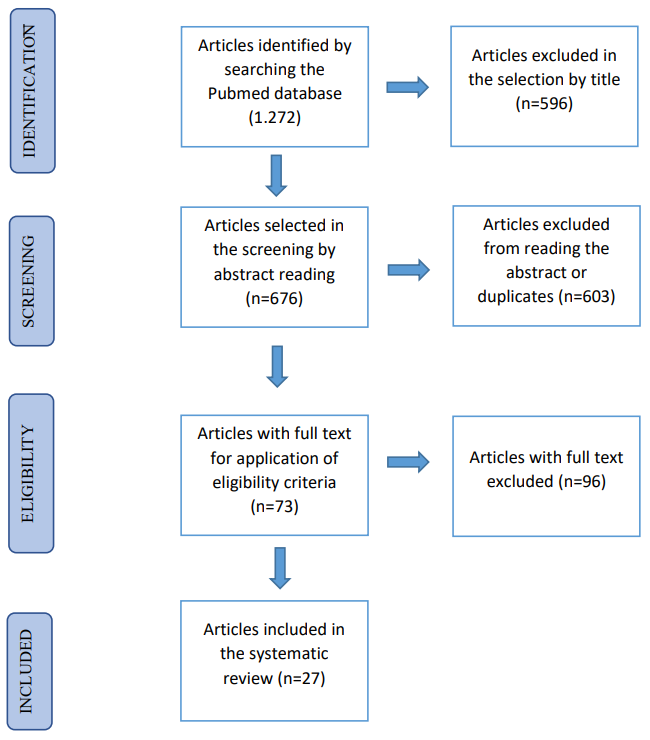
This systematic review identifies and discusses the mechanisms of 27 studies Table 1 reporting the association between pesticide exposure and immune system dysregulation. Only ten (10) studies addressed the relationship with cancer (10/27). Furthermore, of the 27 studies, seven (7) were conducted in humans, nine (9) in experimental animals, and 11 in vitro studies involving cell cultures. As for the countries where the studies were conducted, the sequence is as follows: United States of America (9/27); Brazil (4/27); Italy (3/27); France and Japan (2/27); and Germany, China, Canada, Iran, Mexico, Poland, and India (1/27).
Human Studies
This systematic review included 7 studies that assessed the impact of exposure to pesticides on the immune system of humans and its relationship with cancer. Sample sizes ranged from 10 Faustini A, et al. [13] to 341 Neta G, et al. [14] individuals, with the age range ranging from newborns to adults. Furthermore, the studies included both sexes and different times of exposure to pesticides from days to years.
Therefore, exposure was analyzed in the studies by at least one indicative factor, and i) 4/7 of the studies analyzed exposure through questionnaires or through work with pesticides (informing whether the person had any type of contact with such substances, regardless of type of exposure), and ii) 3/7 studies analyzed exposure by quantifying pesticide levels in biological samples, 1/3 in urine samples Costa C, et al. [15] and 2/3 in blood samples [14, 16].
| Author and year | Title article | Access link |
|---|---|---|
| Sevan, et al. [6] | Aldicarb suppresses macrophage but not natural killer (NK) cell-mediated cytotoxicity of tumor cells. | DOI: 10.1007/BF01701987 |
| Jacobsen-Pereira, et al. [9] | Immune response of Brazilian farmers exposed to multiple pesticides | DOI: 10.1016/j.ecoenv.2020.110912 |
| Arici, et al. [10] | Does pendimethalin develop in pancreatic cancer induced inflammation? | DOI: 10.1016/j.chemosphere.2020.126644 |
| Camargo, et al. [11] | Pesticide dichorvos induces early solid Ehrlich tumoral development associated with a non-protective pro- inflammatory response | DOI:10.1007/s11356-017-1104-x |
| Faustini, et al. [13] | Immunological Changes among Farmers Exposed to Phenoxy Herbicides: Preliminary Observations | DOI: 10.1136/oem.53.9.583 |
| Neta, et al. [14] | Fetal Exposure to Chlordane and Permethrin Mixtures in Relation to Inflammatory Cytokines and Birth Outcomes | DOI: 10.1021/es103417j |
| Costa, et al. [15] | Cytokine patterns in greenhouse workers occupationally exposed to α-cypermethrin: an observational study | DOI: 10.1016/j.etap.2013.07.004 |
| Volker, et al. [16] | Associations of Dichlorodiphenyltrichloroethane (DDT) 4.4 and Dichlorodiphenyldichloroethylene (DDE) 4.4 Blood Levels with Plasma IL-4 | DOI: 10.1080/00039890209602086 |
| Pizatti, et al. [17] | Toxicoproteomics Disclose Pesticides as Downregulators of TNF-α, IL-1β and Estrogen Receptor Pathways in Breast Cancer Women Chronically Exposed | DOI: 10.3389/fonc.2020.01698 |
| Fenga, et al. [18] | IL-17 and IL-22 serum levels in greenhouse workers exposed to pesticides | DOI: 10.1007/s00011-014-0769-6 |
| Teyssier, et al. [19] | Some Plant Defense Stimulators can induce IL-1β production in human immune cells in vitro | DOI: 10.1016/j.toxrep.2020.02.013 |
| Karrow, et al. [20] | Oral exposure to atrazine modulates cell-mediated immune function and decreases host resistance to the B16F10 tumor model in female B6C3F1 mice | DOI: 10.1016/j.tox.2004.12.002 |
| Zeng, et al. [21] | Hematopoietic stem cell and immunotoxicity in zebrafish embryos induced by exposure to Metalaxyl-M | https://doi.org/10.1016/j. scitotenv.2021.152102 |
| Quinteiro-Filho, et al. [22] | Effect of cyhalothrin on Ehrlich tumor growth and macrophage activity in mice | DOI:10.1590/S0100- 879X2009001000006 |
| Jorsaraei, et al. [23] | Immunotoxicity effects of carbaryl in vivo and in vitro | DOI: 10.1016/j.etap.2014.09.004 |
| Medina-Buelvas, et al. [24] | Genotoxic and immunotoxic effects of the organophosphate metabolite diethyldithiophosphate (DEDTP) in vivo | DOI: 10.1016/j.taap.2019.01.023 |
Table 1: Results in chronological order and publication of studies selected in full to compose the systematic review.
- Noworyta-
- Głowacka, et al.
- [25]
- Influence of chlorpyrifos on the profile of subpopulations of immunoactive cells and their phagocytic activity in an experimental in vivo model https://pubmed.ncbi.nlm.nih. gov/23020043/
- Téllez-Bañuelos, et al. [26]
- Chronic exposure to endosulfan induces inflammation in murine colon via β-catenin expression and IL-6 production
- DOI: 10.1080/1547691X.2016.1206998
- Burow, et al. [27]
- Effects of environmental estrogens on tumor necrosis factor α-mediated apoptosis in MCF-7 cells
- DOI: 10.1093/carcin/20.11.2057
- Udoji, et al. [28]
- Immunosuppressive effects of triclosan, nonylphenol, and
- DDT on human natural killer cells in vitro
- DOI: 10.3109/15476911003667470
- Effects of DDT and triclosan on tumor-cell binding capacity and cell-surface protein expression of human natural killer cells
- DOI: 10.1002/jat.2767
- Hurd-Brown, et al.
- [29]
- Martin, et al. [30]
- The organochlorine pesticides pentachlorophenol and dichlorodiphenyltrichloroethane increase secretion and production of interleukin 6 by human immune cells
- DOI: 10.1016/j.etap.2019.103263
- Whalen, et al. [31]
- Immunomodulation of human natural killer cell cytotoxic function by triazine and carbamate pesticides
- DOI: 10.1016/s0009-2797(03)00027-9
- Bifenthrin insecticide promotes oxidative stress and increases inflammatory mediators in human neuroblastoma cells through NF-kappaB pathway
- Gargouri, et al.
- [32] https://doi.org/10.1016/j. tiv.2020.104792
- Shah, et al. [33]
- Organochlorine pesticides induce inflammation, ROS production, and DNA damage in human epithelial ovary cells: An in vitro study
- DOI: 10.1016/j.chemosphere.2019.125691
- Hurd, et al.
- Pentachlorophenol decreases tumor-cell-binding capacity and cell-surface protein expression of human natural killer cells
- DOI: 10.1002/jat.1781
Table 2: Results in chronological order and publication of studies selected in full to compose the systematic review.
Another important factor evaluated was the frequency with which classes of pesticides were analyzed, with greater homogeneity being observed in two of the seven studies that analyzed patients exposed to mixtures of fungicides, herbicides and insecticides [9, 17]. Three of the seven studies verified insecticides, including pyrethroids [14, 15, 16]. Another study (1/7) analyzed pyrethroids and organophosphates Fenga C, et al. [18], and finally, another (1/7) analyzed herbicides [13].
The immune system was the main primary endpoint analyzed in the studies in question, being evaluated by different methodological parameters, including cells and cytokines from the innate and adaptive response system and it was observed that all classes of pesticides interfere with different immune responses and critical cytokines in carcinogenesis.
Regarding the main findings, Faustini, et al. [13] demonstrated that metabolites of pesticides classified as herbicides have negative regulatory effects on T helper cells and cytotoxic T cells (CTL), causing a reduction of these cell types in the exposed organism, including circulating T helper cells (CD4), suppressor cells (CD8) and natural killer (NK) cells. Thus, the immune system would be compromised and could lead to the process of lymphomagenesis [13].
Furthermore, according to Volker, et al. [16], pesticides, when identified in farmers’ blood, cause cellular suppression of Th1-type cytokines, specifically interleukin 2 (IL-2) and interferon-gamma (IFN-γ), in addition to inducing Th2-type cytokines, such as interleukin 4 (IL-4). These Th2 cytokines inhibit Th1 cytokines and cellular immune responses, which may be related to the emergence of tumor activity. In this study, no findings were reported for any disease or cancer. However, the literature shows that these cytokines are closely associated with tumorigenesis [16].
In this same line of study, Neta, et al. [14] in their study described a positive association of permethrin with lower levels of anti-inflammatory expression of interleukin 12 (IL- 12), in addition to lower levels of interleukin-1β (IL-1β) in the umbilical cord of newborns. They also observed a decrease in a cytokine responsible for mediating inflammation, but without association with cancer [14].
Fenga, et al. [18] investigated exposure to neonicotinoid, pyrethroid and organophosphate pesticides in relation to interleukins 17 (IL-17) and 22 (IL-22) in healthy greenhouse workers without any symptoms or illnesses, all using equipment protection correctly. No interference with the IL-17 response was demonstrated; however, there was stimulation of the IL-22 response. These findings show that pesticide exposure can reduce host defenses against cancer through oxidative stress and impaired Th1 function.
Individuals exposed to the pyrethroid cypermethrin demonstrated changes in cytokines and immune response mediators related to pro-inflammatory interleukins. In addition to decreased antitumor immunity and lower levels of IL-2, IL-8 and IFN-γ, both correlate with tumor growth and shorter survival [15].
Exposures to different grouped classes of pesticides (fungicides, insecticides and herbicides) analyzed by Jacobsen-Pereira, et al. [9] caused a significant increase in the activity of innate immunity cells and the number of total T cells, concomitant with a significant decrease in the total number of B cells (regulatory B cells and plasma cells).
Still on the different classes, a study involving women with breast cancer exposed and not exposed to pesticides, carried out by Pizzatti, et al. [17], once again observed the suppression of Th1 response cytokines through tumor necrosis factor (TNF-α). This study provided us with associations with breast cancer, revealing that pesticides affect the immune system, causing immunosuppression [17].
Only three studies in humans specifically addressed the topic of cancer. Nonetheless, in all studies, cancer was somehow mentioned as an important secondary outcome of immune dysregulation [15, 16, 17, 18].
Animal Studies
Animal studies Pereira CHJ, et al. [9, 19] show a positive association between pesticide and cytokine exposure and immune system dysregulation in models using rats, mice and fish. Furthermore, studies have shown an association of these pesticides with tumor induction, even indirectly.
Karrow, et al. [20], observed in their study an increase in splenic CD8 T cells, cytotoxic T cells and leukocyte response in mice exposed to the pesticide atrazine. It was mainly observed that there was a reduction in host resistance and the formation of tumors in animals exposed to the pesticide.
These findings may be due to early exposure, in which the first defense cells are activated and have not yet suffered immunological wear during a long period of exposure.
Selvan, et al. [6] analyzed the effects of carbamate on immune system cells involved in carcinogenesis in rats, where they observed that this pesticide, even at low doses, could suppress macrophage-mediated tumor cell death without affecting NK cell lysis.
In another study, where the fungicide metalaxyl-M was tested in a zebrafish animal model, it was discovered that embryos exposed to the fungicide showed impaired growth and a significant decrease, dependent on exposure, in the number of neutrophils, macrophages and cells. It’s not great. Furthermore, taking into account that all immune cells are derived from pluripotent stem cells (HSCs), the signaling pathway for HSC production was also studied, and a change was found that resulted in the reduction of these cells, which explains the reduction of cells of the innate immune system [21].
A study that evaluated the action of the insecticide cyhalothrin in mice also observed a decrease in innate immunity cells, through a reduction in the activity of peritoneal macrophages. This study further suggests that these data provide evidence that cyhalothrin simultaneously alters host resistance to the growth of Ehrlich tumor, a mammary carcinoma induced in female mice [22].
Furthermore, the same pesticide mentioned above, exposed to rats, acted to reduce IL-2, IFN-γ, IL-1β and TNF-α, and increased the cytokines IL-4 and IL-10. Furthermore, the pesticide suppressed the functions of lymphocytes and macrophages. Thus, there was an increase in Th2 and polarization in the Th1/Th2 balance, as IFN-γ was reduced and IL-4 and IL-10 increased. This mechanism may contribute to neoplasms through immunotoxicity and imbalance of Th1 and Th2, for which Th1 function has decreased and Th2 function has increased [23].
When exposed to organophosphates, Medina-Buelvas, et al. [24] evaluated the production of pro- and anti- inflammatory cytokines in a mouse model, where they observed an increase in the production of IL-6 and IL-10 and a reduction in CD3 T lymphocytes, concomitant with an increase in CD4 and CD8 T lymphocytes. Therefore, it was observed that exposure to low doses of the pesticide can alter immune system cells and modify the innate response. Furthermore, the same chemical pesticide was tested in the diet of male Wistar rats, which did not show critical statistical results in immune system cells, which can be explained by the exposure time (28 days) [25].
Camargo, et al. [11] also analyzed the exposure of organophosphate pesticide metabolites against immune cells but associated it with cancer. They observed that the insecticide increased the cytokine TNF-α at the beginning of tumor development in exposed mice, even at low doses, inducing death by apoptosis and inflammatory responses.
Finally, when analyzing cytokines in response to endosulfan exposure in rats, Téllez-Bañuelos, et al. [26], as well as other previously reported studies Teyssier L, et al. [9, 19] showed an increase in IL-6. Furthermore, in this study, cytokines and TNF-α were increased.
In vitro Studies
Eleven studies involving in vitro cell culture were selected. The cells were exposed to different pesticides, with the majority of studies using the DDT metabolite, precisely four studies Burow ME, et al. [27, 28, 29, 30], which showed the following results: decreased function of NK cells to bind to tumor cells and lysis of these cells; stimulation of IL-1β and IL-6; decreased cell surface protein CD16 and TNF-α. The possible mechanisms cited by the authors were genetic factors due to increased translation, transcription and stability of messenger RNA [27, 28, 30].
Disorders involving decreased function of NK cells have been associated with the lack of ability of these cells to bind to and lyse tumor cells. These same losses of function have been observed in cells exposed to atrazine and carbamates Whalen MM, et al. [31] pyrethroids Gargouri B, et al. [32] and organochlorines [29, 33].
Overexpression of inflammatory cytokines TNF-α, IL-6 and IL-1β was also found Shah HK, et al. [32, 33]. These findings play a central role in tumor progression [33]. It was observed that herbicides were correlated with pancreatic cancer through the induction of significant oxidative damage but no change in IL-16 and IL-8 levels [10].
One of the studies [19] addressed exposure to plant protection products (PPP), with one of these compounds containing Acibenzolar-S-methyl (ASM) particles, inducing an increase in IL-1β. Another compound containing laminarin strongly induced the expression of this same interleukin, even at a low dose. This result differs from the results in humans, showing that occupational exposure decreases this cytokine and cytokines of the same type of response, Th1.
Discussion
This review presents the mechanisms discussed in the literature of how exposure to pesticides affects the immune system and, consequently, how it can affect carcinogenesis. These findings show deregulation in several cellular and humoral components of the immune response through different mechanisms, which act in the origin, progression or worsening of cancer Figure 2.
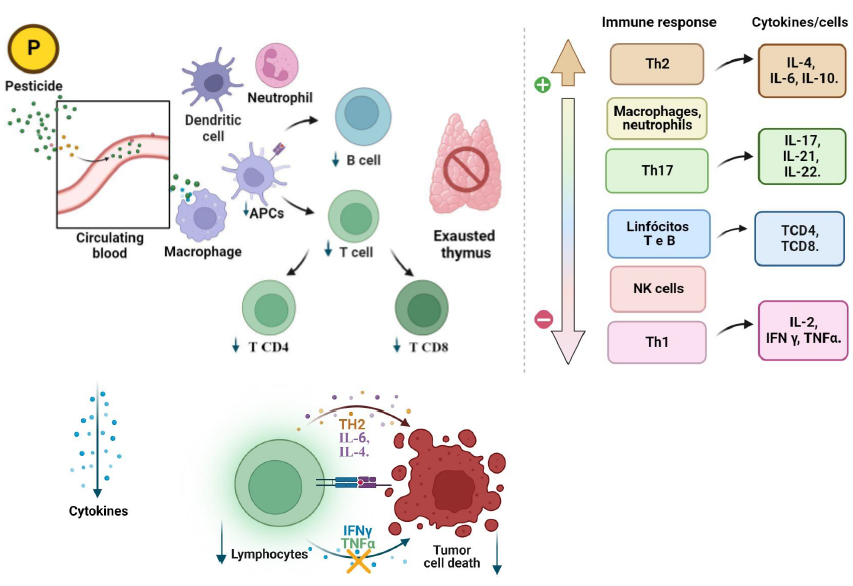
Pesticides enter the bloodstream, being engulfed by Antigen-presenting cells (APCs), dendritic cells, and macrophages. Then, they are forwarded to lymphoid organs such as the thymus, which may become exhausted and diminish cells of the B-lymphocyte, T-lymphocyte, TCD4, and TCD8 types. Due to this mechanism, there is a downregulation of cytokines, increased Th2 response, decreased Th1, and increased tumor susceptibility.
Due to their low molecular weight, pesticides easily penetrate the body and are distributed throughout cells and tissues, breaking the main physical immune barrier (the skin) and exerting their immunotoxicity on both innate and adaptive immunity. These chemicals have a high capacity for bioaccumulation and can act directly on cells or complex proteins in our body, forming haptens (small molecules), easily recognized by the immune system [34].
Chronic exposure, as occurs in the occupational exposure of farmers over several years, can compromise our body’s defense cells, deregulating their function and reducing their number. Thus, other cells try to meet the demand and are overexpressed or repressed, depending on the type of pesticide and the characteristics of exposure, resulting in immunological exhaustion. This depletion reduces the cells that act directly and indirectly on cancer, which can promote growth or even trigger metastasis factors and a worse prognosis, conferring unwanted aggressiveness [17, 20, 35].
With the constant increase in stimuli caused by pesticides, immunological and carcinogenic homeostasis is altered, generating a cascade that deregulates all processes that should occur naturally in the body [24].
TNF-α-producing cells decreased as exposure increased and time lengthened, as in occupational exposure [17, 23]. TNF-α has a multifunctional role in tumor immunity and can induce apoptosis and proliferation of cancer cells, depending on the stimulus. When coming from the tumor cells themselves, TNF-α favors their multiplication, when produced by a response from the immune system, it has the opposite role, helping to fight the tumor and increasing the production of other cytokines that help in this process, which together make part of the Th1 response pattern, this pathway being suitable to act against tumor growth and progression [11, 15, 16, 17, 23, 26, 32].
In relation to carcinogenesis, the ideal would be an organism with these cytokines in high quantities, that is, the opposite of what was proposed by exposure in the studies mentioned. This would occur because these cytokines directly regulate antitumor pathways, indicating to the organism the necessary pathways to stop the tumor, recruiting other cytotoxic cells, such as lymphocytes, or even exacerbating existing cytokines in the environment. This process prevents tumor progression and metastasis to distant organs, providing a better prognosis for patients [7, 36].
Furthermore, studies highlight a reduction of type B cells, both regulatory and plasma B cells, which may be related to dysfunction of T lymphocyte activation or pesticide-induced immunotoxicity of lymphoid organs [9]. Such evidence corroborates the study by Lee, et al. [37, 38], who demonstrated that T and B cells decrease upon exposure to pesticides.
In addition to the decrease in T and B lymphocytes, pesticides affect CD4+ T lymphocytes, CD8+ T lymphocytes and CTL, showing that this is a pathway dependent on activation factors. Thus, when a pathway is altered, there is also a decrease in the secretion of cytokines responsible for signaling the functions or production of other cells. Impairment of a pathway creates a cascade effect and, therefore, immunosuppression, which is favorable to the development of tumors [13, 15, 19]. Studies show us that Th1 and Th17 responses are essential in activating cells and producing cytokines in immunity against tumors, in surveillance against tumor initiation, or even conferring an unfavorable effect. prognosis for cancer patients when compromised [16, 18].
According to the literature, the role of the Th17 response appears adaptive to the environment; depending on the situation, such as in a tumor microenvironment, it can be harmful. However, when activated by the immune system, it helps fight and develop cancer cells [39]. Furthermore, studies show an exacerbated increase in Th2 responses, and this type of response is considered the main promoter of tumors, represented here by the production of interleukins IL-4 and IL-10. Thus, immune dysregulation and the disordered cascade of cytokine production and suppression plus the increase in the Th2 response at the expense of Th1 favor an extremely pro-tumor microenvironment, considerably increasing the risk of cancer in people exposed to pesticides [16, 23].
Finally, it is worth mentioning that the NK cell also showed changes in our review. Studies show that exposure to pesticides causes a reduction in these cells in humans [6, 13] creating a microenvironment favorable to the development of cancer, as they are responsible for recognizing foreign cells and inducing apoptosis, preventing the onset of neoplasia [40].
Conclusion
Given the above, it was possible to verify that exposure to pesticides causes immunological disturbances that affect both innate and adaptive immunity. These disturbances interfere with the functionality of the immune system by deregulating various immune cells and cytokine production, leading to immunosuppression. Such deregulation compromises the body’s ability to effectively defend against infections and other diseases. Additionally, chronic exposure to pesticides can result in an unfavorable prognosis for affected populations due to its significant role in cancer progression. Finally, continuous suppression and poor management of the immune response create an environment conducive to cancer development and exacerbate existing health problems, highlighting the urgent need for better understanding and regulation of pesticide exposure.
Declaration of Conflicts of Interest
The authors declare they have no actual or potential competing financial interests.
References
-
Araujo CAT, Camel TO (2020) Os organofosforados: da segunda guerra mundial à agricultura.
-
Cassal VB, Azevedo LF, Ferreira RP, Silva DG, Simao RS (2014) Pesticides: a review of their consequences for public health. REGET 18(1): 437-445.
-
Araujo AJ, Lima JS, Moreira JC, Jacob SC, Soares MO, et al. (2007) Multiple exposure to pesticides and impacts on health: a cross-section study of 102 rural workers, Nova Friburgo, Rio de Janeiro State, Brazil. Cien Saude Colet 12(1): 115-130.
-
Benedetti D, Nunes E, Sarmento M, Porto C, Santos CEI, et al. (2013) Genetic damage in soybean workers exposed to pesticides: evaluation with the comet and buccal micronucleus cytome assays. Mutat Res 752(1-2): 28-33.
-
Miranda AC, Moreira JC, Carvalho R, Peres F (2007) Neoliberalism, pesticide consumption and food sovereignty crisis in Brazil. Cien Saude Colet 12(1): 7-14.
-
Selvan RS, Dean TN, Misra HP, Nagarkatti PS, Nagarkatti M (1989) Aldicarb suppresses macrophage but not natural killer (NK) cell-mediated cytotoxicity of tumor cells. Bulletin of environmental contamination and toxicology 43: 676-682.
-
Panis C, Kawassaki ACB, Crestani APJ, Pascotto CR, Bortoloti DS, et al. (2022) Evidence on human exposure to pesticides and the occurrence of health hazards in the Brazilian population: a systematic review. Front Public Health 9: 787438.
-
Thong T, Forte CA, Hill EM, Colacino JA (2019) Environmental exposures, stem cells, and cancer. Pharmacol Ther 204: 107398.
-
Pereira CHJ, Cardoso CC, Gehlen TC, Santos CR, Silva MCS (2020) Immune response of Brazilian farmers exposed to multiple pesticides. Ecotoxicol Environ Saf 202: 110912.
-
Arici M, Abudayyak M, Boran T, Ozhan G (2020) Does pendimethalin develop in pancreatic cancer induced inflammation?. Chemosphere 252: 126644.
-
Camargo MR, Akamine PVT, Venturini J, Arruda MSP, Franchi CAS (2018) Pesticide dichorvos induces early solid Ehrlich tumoral development associated with a non-protective pro-inflammatory response. Environ Sci Pollut Res Int 25(8): 7681-7687.
-
Moher D, Shamseer L, Clarke M, Ghersi D, Liberati A, et al. (2015) Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst Rev 4(1): 1.
-
Faustini A, Settimi L, Pacifici R, Fano V, Zuccaro P, et al. (1996) Immunological changes among farmers exposed to phenoxy herbicides: preliminary observations. Occup Environ Med 53(9): 583-585.
-
Neta G, Goldman LR, Barr D, Apelberg BJ, Witter FR, et al. (2011) Fetal exposure to chlordane and permethrin mixtures in relation to inflammatory cytokines and birth outcomes. Environ Sci Technol 45(4): 1680-1687.
-
Costa C, Rapisarda V, Catania S, Di Nola C, Ledda C, et al. (2013) Cytokine patterns in greenhouse workers occupationally exposed to α-cypermethrin: an observational study. Environ Toxicol Pharmacol 36(3): 796-800.
-
Volker D, Wolfgang H, Klausdieter B, Caner S, Christian C, et al. (2010) Associations of Dichlorodiphenyltrichloroethane (DDT) 4.4 and Dichlorodiphenyldichloroethylene (DDE) 4.4 blood levels with plasma IL-4. Arch Environ Health 57(6): 541- 547.
-
Pizzatti L, Kawassaki ACB, Fadel B, Nogueira FCS, Evaristo JAM, et al. (2020) Toxicoproteomics disclose pesticides as downregulators of tnf-α, il-1β and estrogen receptor pathways in breast cancer women chronically exposed. Front Oncol 10: 1698.
-
Fenga C, Gangemi S, Catania S, Luca A, Costa C (2014) IL-17 and IL-22 serum levels in greenhouse workers exposed to pesticides. Inflamm Res 63(11): 895-897.
-
Teyssier L, Sorci G, Chluba J, Aime S, Wendehenne D, et al. (2020) Some Plant Defense Stimulators can induce IL-1β production in human immune cells in vitro. Toxicol Rep 7: 413-420.
-
Karrow NA, McCay JA, Brown RD, Musgrove DL, Guo TL, et al. (2005) Oral exposure to atrazine modulates cell-mediated immune function and decreases host resistance to the B16F10 tumor model in female B6C3F1 mice. Toxicology 209(1): 15-28.
-
Zeng S, Peng Y, Ma J, Ge Y, Huang Y, et al. (2022) Hematopoietic stem cell and immunotoxicity in zebrafish embryos induced by exposure to Metalaxyl-M. Sci Total Environ 809: 152102.
-
Quinteiro-Filho WM, Righi DA, Palermo-Neto J (2009) Effect of cyhalothrin on ehrlich tumor growth and macrophage activity in mice. Braz J Med Biol Res 42: 912-917.
-
Jorsaraei SGA, Maliji G, Azadmehr A, Moghadamnia AA, Faraji AA (2014) Immunotoxicity effects of carbaryl in vivo and in vitro. Environ Toxicol Pharmacol 38(3): 838- 844.
-
Medina-Buelvas D, Estrada-Muniz E, Flores-Valadez M, Vega L (2019) Genotoxic and immunotoxic effects of the organophosphate metabolite diethyldithiophosphate (DEDTP) in Vivo. Toxicol Appl Pharmacol 366: 96-103.
-
Noworyta-Glowacka J, Bankowski R, Siennicka J, Wiadrowska B, Beresinska M, et al. (2011) Influence of chlorpyrifos on the profile of subpopulations of immunoactive cells and their phagocytic activity in an experimental in vivo model. Ann Agric Environ Med 19(3): 483-485.
-
Téllez-Bañuelos MC, Haramati J, Franco Topete K, Peregrina Sandoval J, Franco-Topete R, et al. (2016) Chronic exposure to endosulfan induces inflammation in murine colon via β-catenin expression and IL-6 production. J Immunotoxicol 13(6): 842-849.
-
Burow ME, Tang Y, Collins-Burow BM, Krajewski S, Reed JC, et al. (1999) Effects of environmental estrogens on tumor necrosis factor α-mediated apoptosis in MCF-7 cells. Carcinogenesis 20(11): 2057-2061.
-
Udoji F, Martin T, Etherton R, Whalen MM (2010) Immunosuppressive effects of triclosan, nonylphenol, and DDT on human natural killer cells in vitro. J Immunotoxicol 7(3): 205-212.
-
Hurd-Brown T, Udoji F, Martin T, Whalen MM (2013) Effects of DDT and triclosan on tumor-cell binding capacity and cell-surface protein expression of human natural killer cells. J Appl Toxicol 33(6): 495-502.
-
Martin TJ, Gabure S, Maise J, Snipes S, Peete M, et al. (2019) The organochlorine pesticides pentachlorophenol and dichlorodiphenyltrichloroethane increase secretion and production of interleukin 6 by human immune cells. Environ Toxicol Pharmacol 72: 103263.
-
Whalen MM, Loganathan BG, Yamashita N, Saito T (2003) Immunomodulation of human natural killer cell cytotoxic function by triazine and carbamate pesticides. Chemico-Biological interactions 145(3): 311-319.
-
Gargouri B, Boukholda K, Kumar A, Benazzouz A, Fetoui H, et al. (2020) Bifenthrin insecticide promotes oxidative stress and increases inflammatory mediators in human neuroblastoma cells through NF-kappaB pathway. Toxicology in vitro 65: 104792.
-
Shah HK, Sharma T, Banerjee BD (2020) Organochlorine pesticides induce inflammation, ROS production, and DNA damage in human epithelial ovary cells: an in vitro study. Chemosphere 246: 125691.
-
Peres F, Moreira JC (2003) E veneno ou e remedio? agrotoxicos, saude e ambiente. Fiocruz 9: 384.
-
Gangemi S, Gofita E, Costa C, Teodoro M, Briguglio G, et al. (2016) Occupational and environmental exposure to pesticides and cytokine pathways in chronic diseases (review). International journal of molecular medicine 38(4): 1012-1020.
-
Horssen VR, Hagen TLMT, Eggermont AMM (2006) TNF-α in cancer treatment: molecular insights, antitumor effects, and clinical utility. The oncologist 11(4): 397- 408.
-
Xue H, Lin B, Ni P, Xu H, Huang G (2010) Interleukin-1B and interleukin-1 RN polymorphisms and gastric carcinoma risk: a meta-analysis. Journal of gastroenterology and hepatology 25(10): 1604-1617.
-
Lee GH, Hwang KA, Choi KC (2019) Effects of Fludioxonil on the cell growth and apoptosis in T and B lymphocytes. Biomolecules 9(9): 500.
-
Karpisheh V, Ahmadi M, Abbaszadeh-Goudarzi K, Saray MM, Barshidi A, et al. (2022) The role of Th17 cells in the pathogenesis and treatment of breast cancer. Cancer cell Int 22(1): 108.
-
Hurd T, Walker J, Whalen MM (2012) Pentachlorophenol decreases tumor-cell-binding capacity and cell-surface protein expression of human natural killer cells. Journal of applied toxicology 32(8): 627-634.
- Pattern of Gonadal Hormones in Oral Testosterone-Supplimented Male Wistar Rats with Diabetes-Induced Hypogonadism
- Re-Evaluation of the Genotoxicity of Currently Used Food Dyes in Mouse Multiple Organs Via Continuous Administration by Drinking Using the Comet Assay
- Pharmacogenetics of Type 2 Diabetes Mellitus: Linking Genetic Variability to Drug Efficacy and its Cardiovascular Outcomes
- Exploratory Proteomic Profiling of SARS-CoV-2 Infected THP-1 Macrophages Reveals Alterations in Inflammatory Response and Cellular Metabolism
- Study of Genotoxicity of Hepatocarcinogens in Multiple Organs in Mice by Feeding and Drinking Using the Comet Assay
- Spirulina Polypeptides Inhibit the Growth of Human Lung Tumor (H460) Cells